Você também pode gostar
- Taller OrgánulosDocumento23 páginasTaller OrgánulosJose TôrrësAinda não há avaliações
- KULBERG Catilevers 150-159 TraducciónDocumento10 páginasKULBERG Catilevers 150-159 Traduccióncintya flores armasAinda não há avaliações
- PeroxisomasDocumento23 páginasPeroxisomasMARIA JOSE NUNEZ MACHADOAinda não há avaliações
- Regulación de La Expresión GénicaDocumento67 páginasRegulación de La Expresión GénicaValeria Olmedo0% (1)
- Bioquimica de La SangreDocumento5 páginasBioquimica de La SangreHolman AndersonAinda não há avaliações
- Examenes Complementarios - OrtodonciaDocumento5 páginasExamenes Complementarios - OrtodonciaPaco EspinozaAinda não há avaliações
- Fases de Remodelación Ósea ApunteDocumento4 páginasFases de Remodelación Ósea ApunteBore LsbbAinda não há avaliações
- Edad DentariaDocumento8 páginasEdad DentariaAlejandro RipepiAinda não há avaliações
- Formato de Ricketts para TareaDocumento5 páginasFormato de Ricketts para TareaAngelica AmadorAinda não há avaliações
- Qué Es Un GenDocumento1 páginaQué Es Un GenIrving Uriel Garcia CarranzaAinda não há avaliações
- Argumentos científicos contra el abortoDocumento9 páginasArgumentos científicos contra el abortoBlake Fhabyan Sanchez BernalAinda não há avaliações
- Movimiento Rectilineo Uniforme (MDocumento14 páginasMovimiento Rectilineo Uniforme (Mfernandoluis0% (1)
- SAPU-practico Informe s12Documento9 páginasSAPU-practico Informe s12Sofia Fabiola Vasquez RamirezAinda não há avaliações
- Calculo de EdadDocumento63 páginasCalculo de EdadJoseCarlos GoicocheaAinda não há avaliações
- Universidad de Cuenca: Caracteristicas Clinicas Y Cefalometricas de Clase IiiDocumento15 páginasUniversidad de Cuenca: Caracteristicas Clinicas Y Cefalometricas de Clase IiiVRam Ma100% (1)
- Celulas MadresDocumento6 páginasCelulas Madresanon_375387773Ainda não há avaliações
- Síndrome de ZellwegerDocumento3 páginasSíndrome de ZellwegerbaironAinda não há avaliações
- Clinica Rubrica Operatoria 2021Documento3 páginasClinica Rubrica Operatoria 2021Emmanuel Bolados ZarzuriAinda não há avaliações
- Practica 4Documento7 páginasPractica 4Amisadai Cruz ReyesAinda não há avaliações
- Protocolo de Cepillado Con OrtodonciaDocumento51 páginasProtocolo de Cepillado Con OrtodonciaDiego LopezAinda não há avaliações
- Biotipos faciales y cefalometríaDocumento7 páginasBiotipos faciales y cefalometríaRomina RossiAinda não há avaliações
- Diferencias de Ancho de Arco Maxilomandibular en Centros de Resistencia Estimados Comparación Entre La Oclusión Normal y La Maloclusión Esquelética de Clase IiiDocumento12 páginasDiferencias de Ancho de Arco Maxilomandibular en Centros de Resistencia Estimados Comparación Entre La Oclusión Normal y La Maloclusión Esquelética de Clase IiiAngeLa PariAinda não há avaliações
- Embriologia Craneofacial DRA RODRIGUEZDocumento8 páginasEmbriologia Craneofacial DRA RODRIGUEZErika Rodriguez MarteAinda não há avaliações
- Modelos Simulacion Numerica en OrtodonciaDocumento35 páginasModelos Simulacion Numerica en OrtodonciaDanny RotzAinda não há avaliações
- TRANSCRIPCIONDocumento63 páginasTRANSCRIPCIONElsa MoranAinda não há avaliações
- Tipos de Cromatina y CromosomasDocumento5 páginasTipos de Cromatina y CromosomasErikaAinda não há avaliações
- Orthodontic Tooth MovementDocumento7 páginasOrthodontic Tooth MovementEli ZaAinda não há avaliações
- Bases Genéticas Del Desarrollo PDFDocumento46 páginasBases Genéticas Del Desarrollo PDFJP Vdlc100% (1)
- Fenómeno de Coactivación Muscular Durante Contracciones Isométricas Máximas PDFDocumento4 páginasFenómeno de Coactivación Muscular Durante Contracciones Isométricas Máximas PDFEzequiel LibradoAinda não há avaliações
- Química OrgánicaDocumento14 páginasQuímica OrgánicaFidel CuestaAinda não há avaliações
- Presion OsmoticaDocumento4 páginasPresion OsmoticaKevin EspínAinda não há avaliações
- Regulación de La Expresión GénicaDocumento17 páginasRegulación de La Expresión GénicaEduardo FeriaAinda não há avaliações
- Guía de Ejercicios - MRU - MRUA - Soluciones PDFDocumento5 páginasGuía de Ejercicios - MRU - MRUA - Soluciones PDFAlexis VegaAinda não há avaliações
- El Aparato Carriere Motion Clase IIDocumento8 páginasEl Aparato Carriere Motion Clase IINatalia Mesa de la CruzAinda não há avaliações
- Átomos ExposiciónDocumento27 páginasÁtomos ExposiciónMiryam Ortiz100% (1)
- GnatofotografíaDocumento11 páginasGnatofotografíaJosé Carlos DávilaAinda não há avaliações
- Recorte Libro Tejido EpitelialDocumento45 páginasRecorte Libro Tejido EpitelialKevin LugoAinda não há avaliações
- Fenomenos BiofisicosDocumento7 páginasFenomenos BiofisicosAngeline Cerezo EscobarAinda não há avaliações
- BiofisicaDocumento8 páginasBiofisicaRober Cruz AraujoAinda não há avaliações
- Cromosomas humanos y cromosomopatíasDocumento27 páginasCromosomas humanos y cromosomopatíasYamir Huarancca ÑaupariAinda não há avaliações
- Mitocondrias y peroxisomasDocumento24 páginasMitocondrias y peroxisomasDaniel Clavijo100% (1)
- Guia de Estudio de Metabolismo de CarbohidratosDocumento14 páginasGuia de Estudio de Metabolismo de CarbohidratosMelissa MartinezanguloAinda não há avaliações
- Análisis CefalómetroDocumento11 páginasAnálisis CefalómetrocamiloAinda não há avaliações
- Informe Laboratorio 3.2Documento7 páginasInforme Laboratorio 3.2Viny Gavilanes100% (1)
- Guía de Práctica de Laboratorio de Fisiología 1: Membrana CelularDocumento21 páginasGuía de Práctica de Laboratorio de Fisiología 1: Membrana CelularMEDALID SHIOMARA ROJAS ESPEZAAinda não há avaliações
- Verticalizacion de Molares Dr. Gustavo Gregoret ESP 2Documento9 páginasVerticalizacion de Molares Dr. Gustavo Gregoret ESP 2Javier Cajo Vasquez100% (1)
- Centro de GravedadDocumento9 páginasCentro de GravedadAlex Rafael Urbano GarciaAinda não há avaliações
- Miembro InferiorDocumento28 páginasMiembro InferiorRosbeily SuberoAinda não há avaliações
- Estrategia PICO para la búsqueda de evidenciaDocumento1 páginaEstrategia PICO para la búsqueda de evidenciaAleja TroyaAinda não há avaliações
- Fragmentos de OkazakiDocumento5 páginasFragmentos de OkazakiRossario Mejía Calderón0% (1)
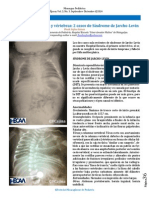
- Síndrome de Jarcho LevinDocumento2 páginasSíndrome de Jarcho Levinnicaragua pediatricaAinda não há avaliações
- La herencia poligénica y las huellas dactilares únicasDocumento5 páginasLa herencia poligénica y las huellas dactilares únicaskaren perezAinda não há avaliações
- Higiene Postural y de La ErgonomíaDocumento17 páginasHigiene Postural y de La ErgonomíaPaola Cristina Gonzalez RoblesAinda não há avaliações
- Propagación rectilínea de la luz: experimento visualDocumento6 páginasPropagación rectilínea de la luz: experimento visualShot Lethal100% (1)
- 23 B. Teórico 2. Estructura Tridimensional de Proteínas. PlegamientoDocumento94 páginas23 B. Teórico 2. Estructura Tridimensional de Proteínas. PlegamientoDebora VassalloAinda não há avaliações
- Las Prestaciones Laborales Obligatorias en México Conforme A La Ley Federal Del TrabajoDocumento4 páginasLas Prestaciones Laborales Obligatorias en México Conforme A La Ley Federal Del TrabajoRuben PeñaAinda não há avaliações
- Presión osmótica y proteínasDocumento14 páginasPresión osmótica y proteínaswendyAinda não há avaliações
- Cues Ti Ones de BioquímicaDocumento2 páginasCues Ti Ones de Bioquímicaapi-26645842Ainda não há avaliações
- Biología celular: Estructura y funciones del retículo endoplasmáticoDocumento9 páginasBiología celular: Estructura y funciones del retículo endoplasmáticoIrene Pulido SerranoAinda não há avaliações
- Distribución y Transporte de Proteínas PDFDocumento38 páginasDistribución y Transporte de Proteínas PDFtaurito26100% (1)
- Procedimientos ExperimentalesDocumento10 páginasProcedimientos ExperimentalesIvvi VarchavskyAinda não há avaliações
- Transcripción en EucariotasssDocumento20 páginasTranscripción en EucariotasssIvvi VarchavskyAinda não há avaliações
- Niveles de Regulación y ModificaciónDocumento6 páginasNiveles de Regulación y ModificaciónIvvi VarchavskyAinda não há avaliações
- Segundo Parcial de BiologiaDocumento31 páginasSegundo Parcial de BiologiaIvvi VarchavskyAinda não há avaliações
- Super BiologiaDocumento14 páginasSuper BiologiaIvvi VarchavskyAinda não há avaliações
- 11 TraducciónDocumento7 páginas11 TraducciónIvvi VarchavskyAinda não há avaliações
- Transcripción en ProcariotasDocumento11 páginasTranscripción en ProcariotasIvvi VarchavskyAinda não há avaliações
- RECUPERATORIO GenéticaDocumento7 páginasRECUPERATORIO GenéticaIvvi VarchavskyAinda não há avaliações
- AnimalesDocumento2 páginasAnimalesIvvi VarchavskyAinda não há avaliações
- Parcialito Animales 1Documento1 páginaParcialito Animales 1Ivvi VarchavskyAinda não há avaliações
- Segundo Parcial de BiologiaDocumento50 páginasSegundo Parcial de BiologiaIvvi VarchavskyAinda não há avaliações
- Arbol AnimalesDocumento1 páginaArbol AnimalesIvvi VarchavskyAinda não há avaliações
- Moléculas OrgánicasDocumento1 páginaMoléculas OrgánicasIvvi VarchavskyAinda não há avaliações
- PROTISTIASDocumento1 páginaPROTISTIASIvvi VarchavskyAinda não há avaliações
- Metabolismo CelularDocumento5 páginasMetabolismo CelularIvvi VarchavskyAinda não há avaliações
- Los Estudiados 2Documento4 páginasLos Estudiados 2Ivvi VarchavskyAinda não há avaliações
- HONGOSDocumento2 páginasHONGOSIvvi VarchavskyAinda não há avaliações
- MICOSDocumento2 páginasMICOSIvvi VarchavskyAinda não há avaliações
- Cuantificación de Adn y Electroforesis en Gel de AgarosaDocumento1 páginaCuantificación de Adn y Electroforesis en Gel de AgarosaIvvi VarchavskyAinda não há avaliações
- Medios de CultivoDocumento2 páginasMedios de CultivoIvvi VarchavskyAinda não há avaliações
- Los EstudiadosDocumento2 páginasLos EstudiadosIvvi VarchavskyAinda não há avaliações
- tp2 EticaDocumento2 páginastp2 EticaIvvi VarchavskyAinda não há avaliações
- Trabajo Práctico 4 y 5 PDocumento10 páginasTrabajo Práctico 4 y 5 PIvvi VarchavskyAinda não há avaliações
- Los Estudiados 3Documento5 páginasLos Estudiados 3Ivvi VarchavskyAinda não há avaliações
- Aislamiento e Identificación de MicroorganismosDocumento3 páginasAislamiento e Identificación de MicroorganismosIvvi VarchavskyAinda não há avaliações
- Ibcm Unq TP 2 y 3Documento11 páginasIbcm Unq TP 2 y 3Ivvi VarchavskyAinda não há avaliações
- TP 1Documento4 páginasTP 1Ivvi VarchavskyAinda não há avaliações
- Cromatografía de GasesDocumento4 páginasCromatografía de GasesIvvi VarchavskyAinda não há avaliações
- Aislamiento Del AdnDocumento2 páginasAislamiento Del AdnIvvi VarchavskyAinda não há avaliações
- RibosomasDocumento6 páginasRibosomasismael coronelAinda não há avaliações
- 3er EFO ChoiceDocumento5 páginas3er EFO ChoiceSer leAinda não há avaliações
- MFH I Semana 6. Orientac Estudio IndependienteDocumento9 páginasMFH I Semana 6. Orientac Estudio IndependienteBranni ParesAinda não há avaliações
- Unidad 2 BiologiaDocumento26 páginasUnidad 2 BiologiaLuna WeckAinda não há avaliações
- Cuales Son Las Etapas Del Proceso de TraduccionDocumento10 páginasCuales Son Las Etapas Del Proceso de TraduccionLurbin J. RomeroAinda não há avaliações
- Tarea Del Laboratorio de BiologiaDocumento2 páginasTarea Del Laboratorio de BiologiaStephany ReyesAinda não há avaliações
- Cuadernillo de Ciencias Naturales Saber-11Documento28 páginasCuadernillo de Ciencias Naturales Saber-11maria fernandaAinda não há avaliações
- Ciencias NaturalesDocumento6 páginasCiencias NaturalesNarvaez Nair EliasAinda não há avaliações
- EXAMEN DE REPOSICION DE NOTA MAS BAJA BIOMOLECULAS - Revisión Del IntentoDocumento17 páginasEXAMEN DE REPOSICION DE NOTA MAS BAJA BIOMOLECULAS - Revisión Del IntentoFabio Jareh ValladaresAinda não há avaliações
- ADN y ARN: estructura y funcionesDocumento22 páginasADN y ARN: estructura y funcionesLuna Hernández Adamaris MonserratAinda não há avaliações
- Apuntes de ABP1 FINALDocumento19 páginasApuntes de ABP1 FINALane.alonsoarrietaAinda não há avaliações
- Síntesis proteínas: Transcripción y traducciónDocumento20 páginasSíntesis proteínas: Transcripción y traducciónDaniel AgudeloAinda não há avaliações
- Diferencias entre eucariota y procariotaDocumento7 páginasDiferencias entre eucariota y procariotaana paola ricalde perazaAinda não há avaliações
- Cuestionario AcDocumento9 páginasCuestionario AcVirginia De La CruzAinda não há avaliações
- 11.naturaleza Molecular Del Gen y Del GenomaDocumento63 páginas11.naturaleza Molecular Del Gen y Del GenomaCristinaOlivaSerranoAinda não há avaliações
- Traduccion Del Adn .Documento1 páginaTraduccion Del Adn .Anlly LuceroAinda não há avaliações
- Genetica MolecularDocumento2 páginasGenetica MolecularnatrixmAinda não há avaliações
- Tipos de Replicación: ¿Por Qué Es Importante La Replicación en El Adn?Documento3 páginasTipos de Replicación: ¿Por Qué Es Importante La Replicación en El Adn?ayleen velarde espinozaAinda não há avaliações
- Bioquímica - Cba-007 - 2022-1 - V2-4Documento11 páginasBioquímica - Cba-007 - 2022-1 - V2-4luciana soto ubilúsAinda não há avaliações
- Antibioticos Mecanismos de AccionDocumento3 páginasAntibioticos Mecanismos de AccionAlexa MoralesAinda não há avaliações
- Ácidos NucléicosDocumento11 páginasÁcidos NucléicosJavier SosaAinda não há avaliações
- Proyecto Final Farmacología Nahabi Itza MartesDocumento165 páginasProyecto Final Farmacología Nahabi Itza MartesNahaibi ArceoAinda não há avaliações
- Tema 6. - Ácidos NucleicosDocumento55 páginasTema 6. - Ácidos Nucleicosmiriamc7realAinda não há avaliações
- Ensayo MetabolismoDocumento3 páginasEnsayo MetabolismoBrianda L Martinez MassAinda não há avaliações
- Nucleótidos y ácidos nucleicosDocumento14 páginasNucleótidos y ácidos nucleicosLesly Maribel Rodríguez Pernillo100% (1)
- TraducciónDocumento6 páginasTraducciónvaleriasantoyo289Ainda não há avaliações
- Teorias Del EnvejecimientoDocumento10 páginasTeorias Del EnvejecimientoSoledadOlivaresAinda não há avaliações
- Montessori - Biologia - 3er Año - Primer Lapso - Formulario de Ejercicios de Genética MolecularDocumento6 páginasMontessori - Biologia - 3er Año - Primer Lapso - Formulario de Ejercicios de Genética MolecularEDUARDO BORREGO100% (1)
- Papel de La Seleniocisteina y de Las SelenioproteínasDocumento14 páginasPapel de La Seleniocisteina y de Las SelenioproteínasinesAinda não há avaliações
- 4° Todos Biología - U1 - Sem 18-05Documento4 páginas4° Todos Biología - U1 - Sem 18-05nicolasAinda não há avaliações