Você também pode gostar
- DN Cross Cutting IssuesDocumento22 páginasDN Cross Cutting Issuesfatmama7031Ainda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Chapter 1 To 5 For Printing.2Documento86 páginasChapter 1 To 5 For Printing.2Senku ishigamiAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Functional DesignDocumento17 páginasFunctional DesignRajivSharmaAinda não há avaliações
- Test Your Knowledge - Study Session 1Documento4 páginasTest Your Knowledge - Study Session 1My KhanhAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Agnes de MilleDocumento3 páginasAgnes de MilleMarie-Maxence De RouckAinda não há avaliações
- Corrosion Fatigue Phenomena Learned From Failure AnalysisDocumento10 páginasCorrosion Fatigue Phenomena Learned From Failure AnalysisDavid Jose Velandia MunozAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- How Drugs Work - Basic Pharmacology For Healthcare ProfessionalsDocumento19 páginasHow Drugs Work - Basic Pharmacology For Healthcare ProfessionalsSebastián Pérez GuerraAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Crown WF-3000 1.2Documento5 páginasCrown WF-3000 1.2Qirat KhanAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Stress-Strain Modelfor Grade275 Reinforcingsteel With Cyclic LoadingDocumento9 páginasStress-Strain Modelfor Grade275 Reinforcingsteel With Cyclic LoadingRory Cristian Cordero RojoAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Parliament of India: Rajya SabhaDocumento64 páginasParliament of India: Rajya SabhaSivapothuraju KonathalaAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Kahneman & Tversky Origin of Behavioural EconomicsDocumento25 páginasKahneman & Tversky Origin of Behavioural EconomicsIan Hughes100% (1)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- SMC 2D CADLibrary English 1Documento590 páginasSMC 2D CADLibrary English 1Design IPGEAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- German Atv-Dvwk Rules and StandardsDocumento25 páginasGerman Atv-Dvwk Rules and StandardsMehmet Emre Bastopcu100% (1)
- Outdoor Air Pollution: Sources, Health Effects and SolutionsDocumento20 páginasOutdoor Air Pollution: Sources, Health Effects and SolutionsCamelia RadulescuAinda não há avaliações
- WEB DESIGN WITH AUSTINE-converted-1Documento9 páginasWEB DESIGN WITH AUSTINE-converted-1JayjayAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Carriage RequirementsDocumento63 páginasCarriage RequirementsFred GrosfilerAinda não há avaliações
- CEE Annual Report 2018Documento100 páginasCEE Annual Report 2018BusinessTech100% (1)
- Strucure Design and Multi - Objective Optimization of A Novel NPR Bumber SystemDocumento19 páginasStrucure Design and Multi - Objective Optimization of A Novel NPR Bumber System施元Ainda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Iec TR 61010-3-020-1999Documento76 páginasIec TR 61010-3-020-1999Vasko MandilAinda não há avaliações
- Working With Difficult People Online WorksheetDocumento4 páginasWorking With Difficult People Online WorksheetHugh Fox IIIAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Problem Set-02Documento2 páginasProblem Set-02linn.pa.pa.khaing.2020.2021.fbAinda não há avaliações
- Guidelines For SKPMG2 TSSP - Draft For Consultation 10.10.17Documento5 páginasGuidelines For SKPMG2 TSSP - Draft For Consultation 10.10.17zqhnazAinda não há avaliações
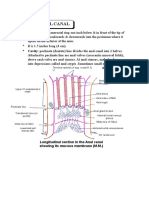
- Anatomy Anal CanalDocumento14 páginasAnatomy Anal CanalBela Ronaldoe100% (1)
- Networker Performance Tuning PDFDocumento49 páginasNetworker Performance Tuning PDFHarry SharmaAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Homework Song FunnyDocumento5 páginasThe Homework Song Funnyers57e8s100% (1)
- Waterstop TechnologyDocumento69 páginasWaterstop TechnologygertjaniAinda não há avaliações
- of Thesis ProjectDocumento2 páginasof Thesis ProjectmoonAinda não há avaliações
- Rule 113 114Documento7 páginasRule 113 114Shaila GonzalesAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Famous Russian PianoDocumento10 páginasFamous Russian PianoClara-Schumann-198550% (2)
- Syllabus DresserDocumento2 páginasSyllabus DresserVikash Aggarwal50% (2)