Você também pode gostar
- Retrovirus (VIH) Principios de Microbiologia Murray 7Documento8 páginasRetrovirus (VIH) Principios de Microbiologia Murray 7Daniel GarciaAinda não há avaliações
- Enfrentando el COVID-19 con los beneficios del cannabis medicinalNo EverandEnfrentando el COVID-19 con los beneficios del cannabis medicinalAinda não há avaliações
- Virus RabiaDocumento36 páginasVirus RabiaMarcelo RealAinda não há avaliações
- 1.VIH-Etiopatogenia de La Infección Por VIHDocumento36 páginas1.VIH-Etiopatogenia de La Infección Por VIHCésar Ariel Rodríguez RavestAinda não há avaliações
- Reseña Sobre El VihDocumento8 páginasReseña Sobre El VihFrancisco Noe hernández JaimeAinda não há avaliações
- Virología Del VIH PDFDocumento32 páginasVirología Del VIH PDFYamil ReyesAinda não há avaliações
- TareahepatitisDocumento3 páginasTareahepatitisAlexandra Stephanie Santa Cruz BellidoAinda não há avaliações
- Mod II-01 Texto Virologia 2021-2022Documento33 páginasMod II-01 Texto Virologia 2021-2022Stephanie Orozco CastilloAinda não há avaliações
- VIH, Resumen FinalDocumento10 páginasVIH, Resumen FinalEduardo MiramontesAinda não há avaliações
- Vih MicrobiologiaDocumento43 páginasVih MicrobiologiaEdgar Modesto JimenezAinda não há avaliações
- Monografía HTLV - GCastro PDFDocumento41 páginasMonografía HTLV - GCastro PDFAldo Caceres Ruiz Diaz100% (1)
- VIHDocumento8 páginasVIHJosé Bancalero de los ReyesAinda não há avaliações
- VIH 2016 MicroDocumento24 páginasVIH 2016 MicroRicardo AguileraAinda não há avaliações
- Tema 20 - VihDocumento8 páginasTema 20 - VihJuan MaldonadoAinda não há avaliações
- Virus de La Inmunodeficiencia HumanaDocumento17 páginasVirus de La Inmunodeficiencia HumanaEdwin J. EscolasticoAinda não há avaliações
- Biologia Molecular Del VihDocumento12 páginasBiologia Molecular Del VihZayana LlerenaAinda não há avaliações
- Tema 20 Retrovirus, Lentivirus y VIHDocumento73 páginasTema 20 Retrovirus, Lentivirus y VIHIsabellaVillalobosAinda não há avaliações
- Resumen RETROVIRUSDocumento21 páginasResumen RETROVIRUSIris Azucena Fernández SalazarAinda não há avaliações
- Esramos, EL TROPISMO DEL VIH Y SU FENOTIPIFICACIÓNDocumento8 páginasEsramos, EL TROPISMO DEL VIH Y SU FENOTIPIFICACIÓNJesabel CarretoAinda não há avaliações
- Virus de La Inmunodeficiencia HumanaDocumento18 páginasVirus de La Inmunodeficiencia HumanaMilton Cordova RomeroAinda não há avaliações
- Inmunopatologgia Del SIDADocumento15 páginasInmunopatologgia Del SIDAsara lopezAinda não há avaliações
- Estructura Del HivDocumento15 páginasEstructura Del HivPolSejLop LópezAinda não há avaliações
- La Estructura Del Virus de VIH Sida. Obra de Arte de La Naturaleza.Documento4 páginasLa Estructura Del Virus de VIH Sida. Obra de Arte de La Naturaleza.Dr. MSc. Byron Núñez Freile100% (3)
- M9T2 - InfecciosasDocumento10 páginasM9T2 - Infecciosasdaniel2056Ainda não há avaliações
- Virus de La Rabia BovinaDocumento19 páginasVirus de La Rabia BovinaAndre Suarez FarfanAinda não há avaliações
- Agente Etiológico VihDocumento4 páginasAgente Etiológico VihKaren VeraAinda não há avaliações
- Anestesia e Infección Por HIVDocumento27 páginasAnestesia e Infección Por HIVmalusetti100% (20)
- Protein ADocumento22 páginasProtein AD̳̿p̳̿ G̶r̶o̶N̶e̶ C-bAinda não há avaliações
- Copia de Pract1-Ej2-8020-abnerDocumento10 páginasCopia de Pract1-Ej2-8020-abnerAbner Pedrero ElíasAinda não há avaliações
- Características Principales Del Genoma VIHDocumento4 páginasCaracterísticas Principales Del Genoma VIHRene VelasquezAinda não há avaliações
- El Virus de La Inmunodeficiencia HumanaDocumento31 páginasEl Virus de La Inmunodeficiencia HumanaRosanna LebronAinda não há avaliações
- Sida y Meiosis y MitosisDocumento16 páginasSida y Meiosis y MitosisdanyAinda não há avaliações
- Tema 5. Familia RetroviridaeDocumento9 páginasTema 5. Familia RetroviridaeYenifer Mejia SuárezAinda não há avaliações
- Inmunoligia Del VihDocumento20 páginasInmunoligia Del VihurpiAinda não há avaliações
- ÉBOLADocumento5 páginasÉBOLANavira TelloAinda não há avaliações
- Cuestionario - Inmunopatogenia Del VIHDocumento4 páginasCuestionario - Inmunopatogenia Del VIHAlbertAinda não há avaliações
- Antagonistas CCR5 Infeccion VIH PDFDocumento8 páginasAntagonistas CCR5 Infeccion VIH PDFSelvin CarballoAinda não há avaliações
- Taller Articulo Covid 19 - MicroDocumento3 páginasTaller Articulo Covid 19 - MicroDavid FrancoAinda não há avaliações
- VIHDocumento19 páginasVIHandres barretoAinda não há avaliações
- HTLV I y IIDocumento22 páginasHTLV I y IIJerson Daryl Zapata GrilloAinda não há avaliações
- Informe VIH & Serologia - Elonis TorresDocumento9 páginasInforme VIH & Serologia - Elonis TorresCYBER CONNECTEDAinda não há avaliações
- Caso VihDocumento20 páginasCaso VihAndres David SanchezAinda não há avaliações
- INMUNOGENETICADocumento78 páginasINMUNOGENETICAMiguel VasquezAinda não há avaliações
- Revision Bibliográfica VEBDocumento18 páginasRevision Bibliográfica VEBasanmiguelhAinda não há avaliações
- TERCERRESUMENDocumento23 páginasTERCERRESUMENOrfelin Daniela PeraltaAinda não há avaliações
- Ayala Tomalá Lorena g10 - VihDocumento7 páginasAyala Tomalá Lorena g10 - VihLorena AyalaAinda não há avaliações
- Estructura y Genoma Del VIHDocumento1 páginaEstructura y Genoma Del VIHEloisa EstrellaAinda não há avaliações
- Virologia: Universidad de Guayaquil Facultad de Ciencias Médicas Carrera de MedicinaDocumento28 páginasVirologia: Universidad de Guayaquil Facultad de Ciencias Médicas Carrera de MedicinaBigi MercuryAinda não há avaliações
- RetrovirusDocumento3 páginasRetrovirusJoa Lisbeth Macias MaciasAinda não há avaliações
- Hepatitis Virales Grupo 2Documento11 páginasHepatitis Virales Grupo 2Valeria Leon AbadAinda não há avaliações
- VIHDocumento13 páginasVIHMaria Paz AndradeAinda não há avaliações
- Protocolo Sars Cov 2Documento5 páginasProtocolo Sars Cov 2FRIDA GUADALUPE HERNANDEZ HERNANDEZAinda não há avaliações
- Virus de La Inmunodeficiencia HumanaDocumento24 páginasVirus de La Inmunodeficiencia HumanaCarlos BustamanteAinda não há avaliações
- Ecob, Estudio de Genetica Monica Manzanares2Documento25 páginasEcob, Estudio de Genetica Monica Manzanares2andresAinda não há avaliações
- Seminario 16: Hiv Y Rotavirus: Hiv - Virus de La Inmunodeficiencia HumanaDocumento8 páginasSeminario 16: Hiv Y Rotavirus: Hiv - Virus de La Inmunodeficiencia Humanajuanpabloseg95Ainda não há avaliações
- Capítulo Herpes Virus PDF 1Documento17 páginasCapítulo Herpes Virus PDF 1Ana valeria Muñoz100% (1)
- Inmunología Kuby PreguntasDocumento14 páginasInmunología Kuby PreguntasCarmen TreviñoAinda não há avaliações
- Herpes VirusDocumento6 páginasHerpes Virussteff23ramosferAinda não há avaliações
- Tarea 9Documento8 páginasTarea 9steve edwinAinda não há avaliações
- Elibro Los Mandalas de Las Flores de BachDocumento44 páginasElibro Los Mandalas de Las Flores de BachElena100% (7)
- Si No Te Gusta Tu Vida Cambia La PDFDocumento80 páginasSi No Te Gusta Tu Vida Cambia La PDFJackyCBAinda não há avaliações
- Manual de Sanacion Por ArqueotiposDocumento104 páginasManual de Sanacion Por Arqueotiposwine4093% (168)
- Autodisciplina Diaria - Habitos - Martin MeadowsDocumento39 páginasAutodisciplina Diaria - Habitos - Martin MeadowsJackyCBAinda não há avaliações
- BIODESCOFIDICACIONDocumento46 páginasBIODESCOFIDICACIONarshia34100% (17)
- Yolibre Sergi TorresDocumento68 páginasYolibre Sergi TorresmiguelangAinda não há avaliações
- El Yaciente Ii PDFDocumento228 páginasEl Yaciente Ii PDFJackyCBAinda não há avaliações
- 17 Reglas Del ExitoDocumento11 páginas17 Reglas Del ExitoEduard Rojas Vaca100% (1)
- El Yaciente IIDocumento228 páginasEl Yaciente IIJackyCBAinda não há avaliações
- Diccionario2016 PDFDocumento721 páginasDiccionario2016 PDFjuankr64100% (10)
- Si No Te Gusta Tu Vida Cambia LaDocumento80 páginasSi No Te Gusta Tu Vida Cambia LaJackyCB100% (1)
- Yolibre Sergi TorresDocumento68 páginasYolibre Sergi TorresmiguelangAinda não há avaliações
- Tarot de Los Bohemios PDFDocumento199 páginasTarot de Los Bohemios PDFfabiano_silva6061Ainda não há avaliações
- Banzhaf Hajo - La Simbologia Y El Significado de Los NumerosDocumento260 páginasBanzhaf Hajo - La Simbologia Y El Significado de Los NumerosRodolfoDeza100% (4)
- Wong Kiew Kit - El Gran Libro de La Medicina ChinaDocumento332 páginasWong Kiew Kit - El Gran Libro de La Medicina Chinaliilu96% (26)
- Stanley Alder-En Busqueda Del Tercer OjoDocumento189 páginasStanley Alder-En Busqueda Del Tercer OjoJackyCB75% (8)
- Boc Victor - El Potencial Del Pensamiento PositivoDocumento14 páginasBoc Victor - El Potencial Del Pensamiento PositivoIvor AlbanecichAinda não há avaliações
- Constitucion Circulo GO Uruguay 1882Documento48 páginasConstitucion Circulo GO Uruguay 1882JackyCBAinda não há avaliações
- Asensi, Manuel - Crítica, Sabotaje y SubalternidadDocumento21 páginasAsensi, Manuel - Crítica, Sabotaje y SubalternidadLaura ManzAinda não há avaliações
- Monografia, DrogadiccionDocumento2 páginasMonografia, DrogadiccionJackyCBAinda não há avaliações
- John Randoph Price Plan de Prosper Id Ad de 40 Dias PDFDocumento7 páginasJohn Randoph Price Plan de Prosper Id Ad de 40 Dias PDFkyokyo69Ainda não há avaliações
- Clymer - La Filosofía Del FuegoDocumento159 páginasClymer - La Filosofía Del Fuegozadkiel20037309100% (3)
- Medicina China - Una Trama Sin Tejedor PDFDocumento393 páginasMedicina China - Una Trama Sin Tejedor PDFdaniella100% (3)
- Chamalu Cambio de MilenioDocumento116 páginasChamalu Cambio de MilenioEdison Nuñez100% (1)
- Encargos Al Universo-Mohr Barbel PDFDocumento52 páginasEncargos Al Universo-Mohr Barbel PDFgamr790421g82100% (1)
- Muhammad El Mensajero de DiosDocumento82 páginasMuhammad El Mensajero de DiosJackyCBAinda não há avaliações
- Michan, Shaya - Manual NaturistaDocumento67 páginasMichan, Shaya - Manual Naturistabruno15000100% (1)
- Sanacion Por ArquetiposDocumento155 páginasSanacion Por ArquetiposJorge Garcia Siles95% (20)
- Apuntes de AcupunturaDocumento58 páginasApuntes de AcupunturaAlicia MoralesAinda não há avaliações
- Curso Gratis Mobu RadiestesiaDocumento11 páginasCurso Gratis Mobu RadiestesiaJackyCBAinda não há avaliações
- Inmunidad Innata y AdaptativaDocumento6 páginasInmunidad Innata y AdaptativaDamaris LoorAinda não há avaliações
- PROTOZOARIOS InfoDocumento1 páginaPROTOZOARIOS InfoEsmeralda Miguel carreraAinda não há avaliações
- Ppt. Riesgos BiologicosDocumento4 páginasPpt. Riesgos BiologicosDavid VerdesotoAinda não há avaliações
- Anemias Hemoliticas AdquiridasDocumento30 páginasAnemias Hemoliticas Adquiridasdavid anton pazosAinda não há avaliações
- Presentación de ChagasDocumento32 páginasPresentación de Chagasfrug0% (1)
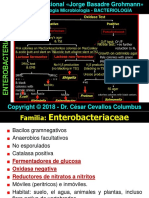
- Familia EnterobacteriumDocumento64 páginasFamilia EnterobacteriumPaolo BarredaAinda não há avaliações
- Diapo VacunasDocumento20 páginasDiapo VacunasIVANARTUROPINOAinda não há avaliações
- 1.sindrome Nefritico - NefroticoDocumento99 páginas1.sindrome Nefritico - Nefroticowanda0612.morianoAinda não há avaliações
- 4-Control de Infecciones en Tuberculosis DR CedillosDocumento34 páginas4-Control de Infecciones en Tuberculosis DR CedillosjoseAinda não há avaliações
- BLASTOMICOSISDocumento15 páginasBLASTOMICOSISCesar Alfredo Cobeñas VillarrealAinda não há avaliações
- Enfermedades Gastrointestinales en MéxicoDocumento16 páginasEnfermedades Gastrointestinales en MéxicoMaria BettyAinda não há avaliações
- VHS 2Documento26 páginasVHS 2Ale VenturaAinda não há avaliações
- Guía Enfermedades Vías Respiratorias SuperioresDocumento3 páginasGuía Enfermedades Vías Respiratorias SuperioresJose Daniel Ardon RuizAinda não há avaliações
- Diarrea Aguda en EquinosDocumento71 páginasDiarrea Aguda en EquinosAngel RuizAinda não há avaliações
- Guia 1 Diluciones InmunologíaDocumento39 páginasGuia 1 Diluciones InmunologíaGissel QuiñonezAinda não há avaliações
- CadenaepidemDocumento27 páginasCadenaepidemSelene Terrones TorresAinda não há avaliações
- Dermatología - BancoAnexo - Con ClavesDocumento3 páginasDermatología - BancoAnexo - Con Clavescal01492Ainda não há avaliações
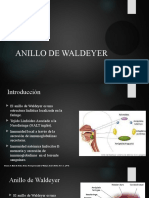
- Anillo WaldeyerDocumento13 páginasAnillo WaldeyerAriadna MéndezAinda não há avaliações
- Coronavirus PPTDocumento26 páginasCoronavirus PPTMeilyn Dayana Principe MendozaAinda não há avaliações
- Historia Natural de La MeningitisDocumento2 páginasHistoria Natural de La MeningitisLiceth BarrosAinda não há avaliações
- Dermatitis SeborreicaDocumento24 páginasDermatitis SeborreicaLizbeth CalleAinda não há avaliações
- ASMA XpoDocumento42 páginasASMA XpoOscar Rujel100% (2)
- ReumatologiaDocumento23 páginasReumatologiaThalia Alaba CastañedaAinda não há avaliações
- ADENITIS CERVICAL ExpoDocumento26 páginasADENITIS CERVICAL ExpoEvelyn Velasquez Gamarra100% (1)
- Bacteriemia en El Paciente CR ́ıticoDocumento10 páginasBacteriemia en El Paciente CR ́ıticoELISAMA HOYOSAinda não há avaliações
- Sifilis CongenitaDocumento22 páginasSifilis CongenitaYolanda Beatriz MirandaAinda não há avaliações
- Historia Clínica N °15Documento24 páginasHistoria Clínica N °15Rishmond Edumar Figueroa Olaguibel100% (2)
- Disertacion TuberculosisDocumento17 páginasDisertacion TuberculosisLeandro CollioAinda não há avaliações
- Anafilaxia FVDocumento24 páginasAnafilaxia FVMaría RaquelAinda não há avaliações
- MAPA CORONO Parte 2 XDDocumento2 páginasMAPA CORONO Parte 2 XDJack H RamosAinda não há avaliações