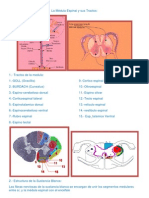
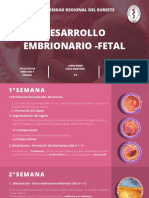
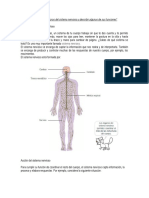
Você também pode gostar
- CITOESQUELETODocumento26 páginasCITOESQUELETOMinerva ShapplinAinda não há avaliações
- Citoesqueleto: FuncionesDocumento14 páginasCitoesqueleto: Funcioneskarina ramos100% (1)
- División CelularDocumento10 páginasDivisión CelularAndrea Fernanda Calderon CuadrosAinda não há avaliações
- Celula HipertonicaDocumento33 páginasCelula HipertonicaErika Viviana MoncadaAinda não há avaliações
- Mitocondria y Respiracion CelularDocumento59 páginasMitocondria y Respiracion CelularMaria RadcliffeAinda não há avaliações
- Taller de Glucolisis y Ciclo de KrebsDocumento7 páginasTaller de Glucolisis y Ciclo de KrebsJhonatan LujanAinda não há avaliações
- Unidad 06 Sistema de EndomembranasDocumento9 páginasUnidad 06 Sistema de EndomembranasjvlpAinda não há avaliações
- Citoesqueleto, ResumenDocumento13 páginasCitoesqueleto, ResumenMKsander GarcíaAinda não há avaliações
- Resumen de Embriologia IvanDocumento97 páginasResumen de Embriologia IvanIvan DarioAinda não há avaliações
- 02 Bioenergetica y TermodinámicaDocumento48 páginas02 Bioenergetica y TermodinámicaJhordy Mamani100% (1)
- Celula Eucariota y ProcariotaDocumento19 páginasCelula Eucariota y ProcariotaChocolatesDoñaMaruAinda não há avaliações
- Introducción de La MitosisDocumento5 páginasIntroducción de La MitosisSneyder Leyton Cárdenas100% (1)
- 8° Guía 3 Organelos CelularesDocumento3 páginas8° Guía 3 Organelos Celularescesar villaAinda não há avaliações
- Sangre Y HematopoyesisDocumento53 páginasSangre Y HematopoyesisAlex Yvan Escobedo Hinostroza100% (1)
- Musculo CardiacoDocumento35 páginasMusculo CardiacoFreddy Cordova Luspa100% (1)
- Aparato CiliarDocumento14 páginasAparato Ciliarlisset8stefany8montoAinda não há avaliações
- Fenómenos Osmóticos en BiologíaDocumento16 páginasFenómenos Osmóticos en BiologíaAna Laura Morales Garcia100% (3)
- EmbriologiaDocumento5 páginasEmbriologiaJhey Kenider Benites IzquierdoAinda não há avaliações
- Reticulo EndoplasmaticoDocumento9 páginasReticulo EndoplasmaticoEnrique Ochoa0% (1)
- Metabolismo Del NitrogenoDocumento43 páginasMetabolismo Del NitrogenoAngelShadow12Ainda não há avaliações
- Insuficiencia Motriz de Origen CerebralDocumento4 páginasInsuficiencia Motriz de Origen Cerebralfisio_end100% (2)
- CentriolosDocumento9 páginasCentriolosANGELA VILLAMILAinda não há avaliações
- Cilios y FlagelosDocumento24 páginasCilios y FlagelosAndres Eleazar ESPINOZA NOLASCO100% (1)
- PPP de NeuroDocumento28 páginasPPP de NeuroJoselyn MuñozAinda não há avaliações
- MICROFILAMENTOSDocumento7 páginasMICROFILAMENTOSalmanza_1973Ainda não há avaliações
- Taller Mitocondria 2019 IDocumento8 páginasTaller Mitocondria 2019 IJESUS ALBERTO SÁNCHEZ GARZONAinda não há avaliações
- Apuntes Tejido MuscularDocumento11 páginasApuntes Tejido Muscularedugalindo8Ainda não há avaliações
- CUELLODocumento48 páginasCUELLOAndrés Rengifo OrjuelaAinda não há avaliações
- Clasificación de Las Células PDFDocumento4 páginasClasificación de Las Células PDFCindy VegaAinda não há avaliações
- Desarrollo Del Aparato RespiratorioDocumento2 páginasDesarrollo Del Aparato RespiratorioAngel Stevens Garcia CallaoAinda não há avaliações
- Proteínas Estructura y FunciónDocumento37 páginasProteínas Estructura y FunciónPaola Duque100% (1)
- La Celula AnimalDocumento9 páginasLa Celula AnimalJuly CollantesAinda não há avaliações
- Ciclo de KrepsDocumento28 páginasCiclo de Krepsmelrc017_906747710Ainda não há avaliações
- Adhesion CelularDocumento30 páginasAdhesion CelularBeto HdzAinda não há avaliações
- Cuestionario CITOESQUELETODocumento2 páginasCuestionario CITOESQUELETOvalentina schalchliAinda não há avaliações
- Desarrollo Del Sistema MuscularDocumento31 páginasDesarrollo Del Sistema MuscularMichelle Cartín TristánAinda não há avaliações
- Potencial de MembranaDocumento17 páginasPotencial de MembranaRobert AcostaAinda não há avaliações
- Trompas de FalopioDocumento21 páginasTrompas de FalopioVeronica Ramirez DamasoAinda não há avaliações
- Texto Académico 1Documento36 páginasTexto Académico 1Patricia Castro SanchezAinda não há avaliações
- Desarrollo Embrionario y FetalDocumento21 páginasDesarrollo Embrionario y FetalBeker solis100% (1)
- Retículo Endoplásmico Pi MZVDocumento29 páginasRetículo Endoplásmico Pi MZVCool BoyLVAinda não há avaliações
- ProtoplasmaDocumento9 páginasProtoplasmaChristopher Guevara CheAinda não há avaliações
- Uniones CelularesDocumento3 páginasUniones CelularesAndreaAinda não há avaliações
- Sistema NerviosoDocumento2 páginasSistema Nerviosooriana alarconAinda não há avaliações
- Membrana CelularDocumento29 páginasMembrana CelularOsvaldo Herrera PalomaresAinda não há avaliações
- Aparato de GolgiDocumento34 páginasAparato de GolgiMoisés J. Duran CanevaroAinda não há avaliações
- Tejido EpitelialDocumento44 páginasTejido EpitelialHenry Petit RodriguezAinda não há avaliações
- APUNTES EmbriologiaDocumento5 páginasAPUNTES Embriologiagabriel ChuquimiaAinda não há avaliações
- Embriologia de Sistema Muscular - Terminado 2Documento14 páginasEmbriologia de Sistema Muscular - Terminado 2Sharito VilelaAinda não há avaliações
- Tejido Muscular PDFDocumento42 páginasTejido Muscular PDFAlex Yvan Escobedo Hinostroza100% (2)
- Ciclo Del Acido Citrico1Documento62 páginasCiclo Del Acido Citrico1Bibiana Andrea PerezAinda não há avaliações
- Guia de Estudio 05 - PseudocelomadosDocumento16 páginasGuia de Estudio 05 - PseudocelomadosRafael Alexander Aparicio MorenoAinda não há avaliações
- La CelulaDocumento29 páginasLa CelulaYanira Soto MedinaAinda não há avaliações
- 3 CelulaDocumento39 páginas3 CelulaAriadna Wolf100% (1)
- Division CelularDocumento7 páginasDivision CelularChristian SumariaAinda não há avaliações
- Organelos No Membranosos - CLASE 3Documento21 páginasOrganelos No Membranosos - CLASE 3Víctor Manuel MoralesAinda não há avaliações
- Citoesqueleto OriginalDocumento16 páginasCitoesqueleto OriginalfranciscoAinda não há avaliações
- CELULADocumento15 páginasCELULADanna Valentina MartinezAinda não há avaliações
- CITOESQUELETODocumento29 páginasCITOESQUELETOElisandra Rodriguez GonzalesAinda não há avaliações
- CitoesqueletoDocumento9 páginasCitoesqueletoOscar RomeroAinda não há avaliações
- Termodinámica QuímicaDocumento22 páginasTermodinámica QuímicaandreaaveAinda não há avaliações
- Membrana CelularDocumento30 páginasMembrana CelularandreaaveAinda não há avaliações
- Organelos CelularesDocumento9 páginasOrganelos CelularesandreaaveAinda não há avaliações
- Edad MediaDocumento6 páginasEdad MediaandreaaveAinda não há avaliações
- Célula y OrganelosDocumento7 páginasCélula y OrganelosandreaaveAinda não há avaliações
- Cascara de YucaDocumento112 páginasCascara de YucaCarlos Perez75% (4)
- Taller 3Documento9 páginasTaller 3Carlos Israel Esparza AndradeAinda não há avaliações
- Hidrolisis de AlmidonDocumento17 páginasHidrolisis de AlmidonPRO BRAYANAinda não há avaliações
- 2do PARCIAL BIOQUÍMICA 2020-2Documento3 páginas2do PARCIAL BIOQUÍMICA 2020-2DANIELA GELVEZAinda não há avaliações
- BIOQUIMICADocumento50 páginasBIOQUIMICANade WentzAinda não há avaliações
- Guiabiomoleculas1medio 120529213729 Phpapp02Documento2 páginasGuiabiomoleculas1medio 120529213729 Phpapp02David AntonioAinda não há avaliações
- Bioquimica - ColagenoDocumento18 páginasBioquimica - ColagenoEstefaniaAinda não há avaliações
- Tarea de Quimica de AluminioDocumento5 páginasTarea de Quimica de AluminioLisbeth MamaniAinda não há avaliações
- Los FanDocumento18 páginasLos FanDayanna Macías ChilánAinda não há avaliações
- Guía de PolímerosDocumento23 páginasGuía de PolímerosVictor Alca SalcedoAinda não há avaliações
- Composicion Quimica Del PescadoDocumento18 páginasComposicion Quimica Del PescadoLuYs CutipaAinda não há avaliações
- 3 Hipótesis Sobre El Origen de La VidaDocumento14 páginas3 Hipótesis Sobre El Origen de La VidaRené CopeticonaAinda não há avaliações
- Efectos Deficiencia ProteinasDocumento11 páginasEfectos Deficiencia ProteinasSofia NiveloAinda não há avaliações
- Desarrollo Evidencia Función de Los AlimentosDocumento4 páginasDesarrollo Evidencia Función de Los AlimentosAngélica ValenciaAinda não há avaliações
- Clase 4 Biomoléculas Orgánicas Proteínas y Ácidos NucleicosDocumento33 páginasClase 4 Biomoléculas Orgánicas Proteínas y Ácidos Nucleicosantonio muñozAinda não há avaliações
- 6019 MP BioquímicaDocumento30 páginas6019 MP BioquímicaLoba NegraAinda não há avaliações
- Determinación Cuantitativa de Urea #4Documento4 páginasDeterminación Cuantitativa de Urea #4Ana0687Ainda não há avaliações
- Dietas MilagrosasDocumento45 páginasDietas MilagrosasKatia Quispe RamirezAinda não há avaliações
- Limpieza Del Cuerpo Desintoxicacion de SistemasDocumento174 páginasLimpieza Del Cuerpo Desintoxicacion de SistemasisidroveAinda não há avaliações
- Key Elemnets Catalogo PDFDocumento7 páginasKey Elemnets Catalogo PDFAndy LombardoAinda não há avaliações
- Artículo CompletoDocumento33 páginasArtículo CompletoValentina RodriguezAinda não há avaliações
- Pau-Genetica-Molecular Soluciones 2016 PDFDocumento12 páginasPau-Genetica-Molecular Soluciones 2016 PDFSilvia MartinezAinda não há avaliações
- Analisis Proximal de Weende y Van SoestDocumento10 páginasAnalisis Proximal de Weende y Van SoestLuz Hadit Calanche Fernandez0% (1)
- Nutrición para FisicoculturistasDocumento27 páginasNutrición para FisicoculturistasEnrique Alexander Hernández PeñalozaAinda não há avaliações
- Aplicaciones de La Biotecnología en El Desarrollo de La Medicina PersonalizadaDocumento8 páginasAplicaciones de La Biotecnología en El Desarrollo de La Medicina Personalizadatrabajos unadAinda não há avaliações
- Tarea N°07 - "Trucha Ahumada y Salado Secado de Pescado"Documento26 páginasTarea N°07 - "Trucha Ahumada y Salado Secado de Pescado"Pamela Ruidias Vélez100% (1)
- Ficha Tecnica ArinaDocumento3 páginasFicha Tecnica ArinaYamii cabezasAinda não há avaliações
- CervezaDocumento48 páginasCervezaStiven OrconAinda não há avaliações
- Interacción HidrofílicaDocumento17 páginasInteracción HidrofílicaJulián BelloAinda não há avaliações
- Alimentación para NadadoresDocumento4 páginasAlimentación para NadadoresWyca WycaAinda não há avaliações