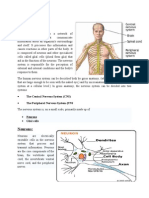
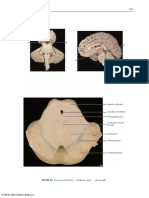
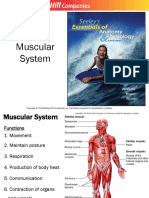
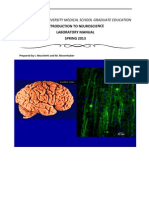
Você também pode gostar
- The Central Nervous SystemDocumento7 páginasThe Central Nervous Systemjoeywap29Ainda não há avaliações
- Kaplan and Sadock's Study Guide and Self-Examination Review in PsychiatryDocumento453 páginasKaplan and Sadock's Study Guide and Self-Examination Review in PsychiatryMizpah Amados100% (14)
- Nervous System LecDocumento26 páginasNervous System LecLuningning Nobleza100% (3)
- The Nervous System - PsychologyDocumento17 páginasThe Nervous System - Psychologyjanet100% (2)
- 2 Human Body Nervous SystemDocumento24 páginas2 Human Body Nervous SystemRain HuntsdaleAinda não há avaliações
- Brain AnatomyDocumento57 páginasBrain AnatomyCarlo BelazaAinda não há avaliações
- Nervous System NotesDocumento6 páginasNervous System NotesAlex Whitwam100% (3)
- (2023) (Stephen Wolfram) - What Is ChatGPT Doing ... and Why Does It Work (Z-Library)Documento123 páginas(2023) (Stephen Wolfram) - What Is ChatGPT Doing ... and Why Does It Work (Z-Library)Lucas OliveiraAinda não há avaliações
- Neurophysiology A Conceptual Approach Carpenter 5th PDFDocumento352 páginasNeurophysiology A Conceptual Approach Carpenter 5th PDFDeepak Kumar100% (7)
- Diploma in Occupational TherapyDocumento47 páginasDiploma in Occupational TherapyAzmi Plus100% (2)
- Reflex PhysiologyDocumento46 páginasReflex Physiologysharmaine_ting100% (1)
- Nervou S System:: NeuronsDocumento16 páginasNervou S System:: NeuronsFizzah100% (4)
- The Nervous System - NotesDocumento12 páginasThe Nervous System - NotesLol lol100% (2)
- Atlas of Functional Neuroanatomy, 3e (Aug 14, 2015) - (146658534X) - (CRC Press)Documento319 páginasAtlas of Functional Neuroanatomy, 3e (Aug 14, 2015) - (146658534X) - (CRC Press)Carmen Ciani100% (9)
- Nervous SystemDocumento20 páginasNervous SystemIsheba Warren100% (3)
- Nervous SystemDocumento46 páginasNervous SystemMac Gab100% (3)
- Action PotentialDocumento53 páginasAction PotentialHusna Amalia100% (2)
- The Muscular SystemDocumento14 páginasThe Muscular SystemOana Maria FrancAinda não há avaliações
- Integumentary System Anatomy and PhysiologyDocumento13 páginasIntegumentary System Anatomy and PhysiologyKBD0% (1)
- Guide Questions AnswersDocumento15 páginasGuide Questions AnswersPrancheska Abigayle Peneyra SantiagoAinda não há avaliações
- 21-Spinal Cord TractsDocumento23 páginas21-Spinal Cord TractsALFAHRUL CAHYADIAinda não há avaliações
- CNS Cross Section QuizDocumento13 páginasCNS Cross Section QuizJay Kay100% (1)
- Nervous SystemDocumento37 páginasNervous SystemShriram100% (2)
- The Nervous System: The Central Processing UnitDocumento85 páginasThe Nervous System: The Central Processing UnitGeoffrey MilesAinda não há avaliações
- The Central Nervous SystemDocumento59 páginasThe Central Nervous SystemJuliville Hora Salinas100% (1)
- Spinal Cord Lesions: Jia Yan-Jie M.D PH.DDocumento170 páginasSpinal Cord Lesions: Jia Yan-Jie M.D PH.Dapi-19916399Ainda não há avaliações
- Lesson 5 - The Powers of The MindDocumento4 páginasLesson 5 - The Powers of The MindCin DyAinda não há avaliações
- PHY 6.11 Motor Reflexes Dr. BartolomeDocumento8 páginasPHY 6.11 Motor Reflexes Dr. BartolomeKate Lynne CamonayanAinda não há avaliações
- Guide Questions AnswersDocumento7 páginasGuide Questions AnswersYda Maxine PalmaAinda não há avaliações
- Seeley's Chapter 8 Nervous SystemDocumento100 páginasSeeley's Chapter 8 Nervous SystemChristine Tapawan89% (9)
- Nervous System - GeneralDocumento84 páginasNervous System - GeneralAman SharmaAinda não há avaliações
- Muscular SystemDocumento117 páginasMuscular SystemAiman Bañaga Arabain100% (1)
- Sensory System: A Tutorial Study Guide: Science Textbook SeriesNo EverandSensory System: A Tutorial Study Guide: Science Textbook SeriesNota: 5 de 5 estrelas5/5 (1)
- Neurophysiology, Neurotransmitters and The Nervous SystemDocumento69 páginasNeurophysiology, Neurotransmitters and The Nervous SystemKUNNAMPALLIL GEJO JOHN100% (2)
- NCM 104 - Nervous SystemDocumento51 páginasNCM 104 - Nervous Systembabeepny807076100% (5)
- Nervous SystemDocumento34 páginasNervous SystemAlmiah Alfarouk100% (1)
- Bluetooth Low Energy in Android Java - Your Guide To Programming The Internet of ThingsDocumento1.100 páginasBluetooth Low Energy in Android Java - Your Guide To Programming The Internet of Thingsde_me30% (1)
- Nervous SystemDocumento32 páginasNervous SystemthwisemanAinda não há avaliações
- A&P Coloring Nervous Worksheet PDFDocumento20 páginasA&P Coloring Nervous Worksheet PDFHelen English100% (1)
- The Nervous System Flashcards - QuizletDocumento11 páginasThe Nervous System Flashcards - QuizletDani AnyikaAinda não há avaliações
- Cranial Nerve 1stDocumento21 páginasCranial Nerve 1stprasun_v100% (1)
- SMP Neuro Lab ManualDocumento37 páginasSMP Neuro Lab Manualsas345sas345Ainda não há avaliações
- Nervous SystemDocumento49 páginasNervous SystemJoyce NieveraAinda não há avaliações
- Frontal Lobe and Temporal Lobe Functions and SyndromesDocumento48 páginasFrontal Lobe and Temporal Lobe Functions and SyndromesShreya Sharma100% (1)
- Overview of Nervous SystemDocumento63 páginasOverview of Nervous SystemMAHARANI100% (1)
- Anatomy of Nervous TissueDocumento54 páginasAnatomy of Nervous TissueNand PrakashAinda não há avaliações
- Autonomic Nervous SystemDocumento41 páginasAutonomic Nervous SystemYanto HaryantoAinda não há avaliações
- Biology G - Lesson 5 - Nervous SystemDocumento59 páginasBiology G - Lesson 5 - Nervous SystemJeremiah Nayosan0% (1)
- Full Download Ebook PDF Human Biology Concepts and Current Issues 8th Edition PDFDocumento41 páginasFull Download Ebook PDF Human Biology Concepts and Current Issues 8th Edition PDFwilliam.decosta187100% (37)
- Autonomic Nervous SystemDocumento55 páginasAutonomic Nervous SystemtiiandiAinda não há avaliações
- Aging Changes in Nervous SystemDocumento333 páginasAging Changes in Nervous SystemHasanErkel100% (2)
- Principles of Neural and Hormonal CommunicationDocumento56 páginasPrinciples of Neural and Hormonal Communicationlynn.dargham2Ainda não há avaliações
- Neurons and SynapseDocumento44 páginasNeurons and Synapsethummiepurple100% (3)
- Complexity A Guided TourDocumento366 páginasComplexity A Guided TourMariana Lira100% (2)
- The Human Nervous SystemDocumento15 páginasThe Human Nervous SystemSiti ZahraAinda não há avaliações
- @MBS MedicalBooksStore 2019 Pain PDFDocumento499 páginas@MBS MedicalBooksStore 2019 Pain PDFRay MaudyAinda não há avaliações
- D.muscular System PDFDocumento8 páginasD.muscular System PDFJohn Lawrence YbanezAinda não há avaliações
- NeurologyDocumento42 páginasNeurologybuzz Q100% (2)
- Behavioral Neuroscience Essentials and Beyond Full ChapterDocumento41 páginasBehavioral Neuroscience Essentials and Beyond Full Chaptercharlie.heiliger836100% (20)
- Anatomy and Physiology of BrainDocumento21 páginasAnatomy and Physiology of BrainNavjot BrarAinda não há avaliações
- Nervous System Flow ChartDocumento1 páginaNervous System Flow Chartstevensvoboda11100% (1)
- Notes On Sensory SystemDocumento4 páginasNotes On Sensory SystemJulienne Sanchez-SalazarAinda não há avaliações
- Anatomy & Physiology 1 Chapter 9 The Nervous System Flashcards - QuizletDocumento7 páginasAnatomy & Physiology 1 Chapter 9 The Nervous System Flashcards - Quizletmalenya1100% (1)
- Students Clinical Case AnalysisDocumento5 páginasStudents Clinical Case Analysisjacc_282Ainda não há avaliações
- Cutaneous Innervation of HeadDocumento4 páginasCutaneous Innervation of HeadAlexandros LazarouAinda não há avaliações
- Nervous SystemDocumento30 páginasNervous SystemkenAinda não há avaliações
- Microsemi FPGA and SoC Products Reliability Report RT0001 v16Documento106 páginasMicrosemi FPGA and SoC Products Reliability Report RT0001 v16de_me3Ainda não há avaliações
- Stop LimitDocumento5 páginasStop Limitde_me3Ainda não há avaliações
- FoCal Target 5r2nsDocumento1 páginaFoCal Target 5r2nsde_me3100% (1)
- Nordic Nrf52840-Dk PBDocumento2 páginasNordic Nrf52840-Dk PBde_me3Ainda não há avaliações
- Gce Marking Scheme: Biology/Human Biology (New) AS/AdvancedDocumento33 páginasGce Marking Scheme: Biology/Human Biology (New) AS/AdvancedRabea2222sweetandsouAinda não há avaliações
- Learning How To Learn Powerful Mental Tools To Help You Master Tough Subjects - Santander IE Online Learning Journey For LearnersDocumento7 páginasLearning How To Learn Powerful Mental Tools To Help You Master Tough Subjects - Santander IE Online Learning Journey For LearnersLukeAinda não há avaliações
- Chapter 20 - MolluscaGastropoda CompressedDocumento27 páginasChapter 20 - MolluscaGastropoda Compressedaulia rahmahAinda não há avaliações
- Learning Objectives - Nervous SystemDocumento3 páginasLearning Objectives - Nervous SystemBelle ChenAinda não há avaliações
- The Irritable Larynx SyndromeDocumento9 páginasThe Irritable Larynx SyndromeLoreto SaavedraAinda não há avaliações
- Flashcards - Topic 14 Coordination and Response - CAIE Biology IGCSEDocumento134 páginasFlashcards - Topic 14 Coordination and Response - CAIE Biology IGCSEGopika KrishnanAinda não há avaliações
- Muscular Nerve Tissue Filll in Notes Student Kd13Documento2 páginasMuscular Nerve Tissue Filll in Notes Student Kd13Greg HayesAinda não há avaliações
- Term Paper AlphaXIIDocumento27 páginasTerm Paper AlphaXIISnehasis MajumdarAinda não há avaliações
- ReflexDocumento2 páginasReflexSarah LewisAinda não há avaliações
- MSC Entrance Examination Topics BiologyDocumento2 páginasMSC Entrance Examination Topics Biologyjohnny brooksAinda não há avaliações
- Neurons Are Not The Only Brain Cells That Think - The EconomistDocumento9 páginasNeurons Are Not The Only Brain Cells That Think - The EconomistdiegocqaAinda não há avaliações
- Course PlanDocumento17 páginasCourse PlanDelphy VargheseAinda não há avaliações
- Johaimah's Report For TNCTDocumento15 páginasJohaimah's Report For TNCTSittiehaya Angnie BAltAinda não há avaliações
- 1.05 Psychobiology: Niels Birbaumer University of Tuèbingen, Germany and Herta Flor Humboldt-University, Berlin, GermanyDocumento58 páginas1.05 Psychobiology: Niels Birbaumer University of Tuèbingen, Germany and Herta Flor Humboldt-University, Berlin, GermanySandra Carolina Valencia LaraAinda não há avaliações
- Chapter 2 NeuronDocumento49 páginasChapter 2 NeuronNithinAinda não há avaliações
- Anes 8 Introduction To Pain ManagementDocumento4 páginasAnes 8 Introduction To Pain ManagementJanica Marie RagsacAinda não há avaliações
- 10 1126@science Aau8302Documento18 páginas10 1126@science Aau8302nicolasAinda não há avaliações
- Reading 3Documento148 páginasReading 3PPP CenterAinda não há avaliações
- Topic 8 - Grey MatterDocumento53 páginasTopic 8 - Grey MatterStudent 365Ainda não há avaliações
- Massachusetts Institute of TechnologyDocumento2 páginasMassachusetts Institute of TechnologyZeynal Abidin ŞabaşAinda não há avaliações
- GenBio 2 MidtermsDocumento9 páginasGenBio 2 MidtermsPanot na BilogAinda não há avaliações
- Musculoskeletal SystemDocumento92 páginasMusculoskeletal SystemBrenda LynAinda não há avaliações
- Biology 12 Chapter 29.2Documento13 páginasBiology 12 Chapter 29.2ismail gamalAinda não há avaliações