Você também pode gostar
- Whales, Dolphins, and Porpoises of the Western North Atlantic: A Guide to Their IdentificationNo EverandWhales, Dolphins, and Porpoises of the Western North Atlantic: A Guide to Their IdentificationAinda não há avaliações
- Longirostris Movement and Behavioral Patterns ToDocumento10 páginasLongirostris Movement and Behavioral Patterns TonanaAinda não há avaliações
- 2010 - Clua Et Al. Behavioural Response of LS To FeedingDocumento10 páginas2010 - Clua Et Al. Behavioural Response of LS To FeedingKaro ZuñigaAinda não há avaliações
- GVI Seychelles Marine Report Jul 2014 - Dec 2015 (Cap Ternay)Documento93 páginasGVI Seychelles Marine Report Jul 2014 - Dec 2015 (Cap Ternay)GVI SeychellesAinda não há avaliações
- Observations of Localised Movements and Residence Times of Wobbegong Sharks (Orectolobus Halei) at Fish Rock, NSW, AustraliaDocumento10 páginasObservations of Localised Movements and Residence Times of Wobbegong Sharks (Orectolobus Halei) at Fish Rock, NSW, AustraliaismailsalaydrusAinda não há avaliações
- tmp3359 TMPDocumento12 páginastmp3359 TMPFrontiersAinda não há avaliações
- Carranza 2006 BiolConservDocumento6 páginasCarranza 2006 BiolConservtortugamarina100% (2)
- 2 HHHJJDocumento13 páginas2 HHHJJFransLei Vadoel SaelAinda não há avaliações
- NOAA Status Report On Coral ReefsDocumento7 páginasNOAA Status Report On Coral ReefsHawaii News NowAinda não há avaliações
- WhaletriviaDocumento4 páginasWhaletriviaapi-348449980Ainda não há avaliações
- Research Expedition To Assess Coral Reef Conditions and Recovery From Mass BleachingDocumento4 páginasResearch Expedition To Assess Coral Reef Conditions and Recovery From Mass Bleachingfranklloyd1975Ainda não há avaliações
- Global Ecology and Conservation: Original Research ArticleDocumento13 páginasGlobal Ecology and Conservation: Original Research ArticleKatie CrolleyAinda não há avaliações
- The Kenya Marine Mammal Network Newsletter. Issue 1Documento5 páginasThe Kenya Marine Mammal Network Newsletter. Issue 1GVI Kenya - Conservation and community developmentAinda não há avaliações
- Population Genetics of Four Heavily Exploited Shark Species Around The Arabian PeninsulaDocumento16 páginasPopulation Genetics of Four Heavily Exploited Shark Species Around The Arabian Peninsulaaljazza03Ainda não há avaliações
- Occurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaDocumento10 páginasOccurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaRizky RamaAinda não há avaliações
- Towards Sustainable Management of The Sardinella Fishery in GhanaDocumento167 páginasTowards Sustainable Management of The Sardinella Fishery in Ghanabrancolee697Ainda não há avaliações
- Decline Dolphines TourismDocumento8 páginasDecline Dolphines TourismArroyo GramajoAinda não há avaliações
- TMP DBA0Documento15 páginasTMP DBA0FrontiersAinda não há avaliações
- Annotated BibliographyDocumento7 páginasAnnotated Bibliographyapi-520649061Ainda não há avaliações
- Small Island Ecosystems: Ekologi Laut Tropis 2015 Tim EkolaDocumento20 páginasSmall Island Ecosystems: Ekologi Laut Tropis 2015 Tim EkolaRIFALDIAinda não há avaliações
- First Report On The Density and Size of Napoleon WrasseDocumento13 páginasFirst Report On The Density and Size of Napoleon WrasseGoresan RimbaAinda não há avaliações
- The Productivity and Public Health Considerations of The Urban Women's Daytime Subsistence Fishery Off Suva Peninsula, Fiji NJ Quinn MT Davis 1997Documento30 páginasThe Productivity and Public Health Considerations of The Urban Women's Daytime Subsistence Fishery Off Suva Peninsula, Fiji NJ Quinn MT Davis 1997Jacque C DiverAinda não há avaliações
- Research Log 3Documento4 páginasResearch Log 3api-510377365Ainda não há avaliações
- Friedlander A Et Al 2005 The State of Coral Reef Ecosystems in NWHIDocumento42 páginasFriedlander A Et Al 2005 The State of Coral Reef Ecosystems in NWHIJuliaParishAinda não há avaliações
- Cephalorhynchus Hectori MauiDocumento4 páginasCephalorhynchus Hectori Mauiapi-637156639Ainda não há avaliações
- Sharks Resource CollectionDocumento2 páginasSharks Resource CollectionJhon de ToledoAinda não há avaliações
- Native Densities, Distribution, and Diurnal Activity of Red Sea Lionfishes (Scorpaenidae)Documento10 páginasNative Densities, Distribution, and Diurnal Activity of Red Sea Lionfishes (Scorpaenidae)Sarah SobhiAinda não há avaliações
- GVI Seychelles: Inside This IssueDocumento7 páginasGVI Seychelles: Inside This IssueGVI SeychellesAinda não há avaliações
- Monk Seal Critical HabitatDocumento1 páginaMonk Seal Critical Habitatapi-290660369Ainda não há avaliações
- Identification and Salinity Tolerance of The Western Hemisphere Mussel MytellaDocumento10 páginasIdentification and Salinity Tolerance of The Western Hemisphere Mussel MytellaNiño Jess Mar MechaAinda não há avaliações
- Whales and Crab FishingDocumento3 páginasWhales and Crab Fishingapi-286094386Ainda não há avaliações
- Ecology Article 2 - Sharks and EcosystemsDocumento3 páginasEcology Article 2 - Sharks and Ecosystemsapi-239443838Ainda não há avaliações
- Molluscs Diversity in Coastal Ecosystem of South Biak, Papua Keanekaragaman Moluska Di Ekosistem Pesisir Biak Selatan, PapuaDocumento13 páginasMolluscs Diversity in Coastal Ecosystem of South Biak, Papua Keanekaragaman Moluska Di Ekosistem Pesisir Biak Selatan, PapuaNur HayatiAinda não há avaliações
- Log 1Documento3 páginasLog 1api-609769059Ainda não há avaliações
- Dolphin Exploitation and Suffering at Se PDFDocumento7 páginasDolphin Exploitation and Suffering at Se PDFBarbaraAinda não há avaliações
- Offshore Dispersion of Ephyrae and Medusae of Aurelia Aurita S.L. (Cnidaria: Scyphozoa) From Port Enclosures: Physical and Biological FactorsDocumento8 páginasOffshore Dispersion of Ephyrae and Medusae of Aurelia Aurita S.L. (Cnidaria: Scyphozoa) From Port Enclosures: Physical and Biological FactorsCintya NevarezAinda não há avaliações
- Adaptive RadiationDocumento4 páginasAdaptive Radiationapi-524269352Ainda não há avaliações
- Wildlife ResearchDocumento7 páginasWildlife ResearchainfebrianAinda não há avaliações
- A Review On Distribution of Crocodylus Porosus (Saltwater Crocodile)Documento7 páginasA Review On Distribution of Crocodylus Porosus (Saltwater Crocodile)NUR ALIAH IZZATI BINTI MOHD RAFI BS20160556Ainda não há avaliações
- Do No So DuttonDocumento13 páginasDo No So DuttontortugamarinaAinda não há avaliações
- tmp3FAE TMPDocumento12 páginastmp3FAE TMPFrontiersAinda não há avaliações
- Lionfish FactsheetDocumento2 páginasLionfish FactsheetugosubaldopiacasseraAinda não há avaliações
- Blue Whale Species ReportDocumento8 páginasBlue Whale Species Reportapi-601591623Ainda não há avaliações
- GVI Seychelles: Inside This IssueDocumento7 páginasGVI Seychelles: Inside This IssueGVI SeychellesAinda não há avaliações
- Rowatetal 2008Documento6 páginasRowatetal 2008Muhammad DanishAinda não há avaliações
- The Great White Shark - EssayDocumento24 páginasThe Great White Shark - EssayDomenica GrangiottiAinda não há avaliações
- Munsch Washington 0250E 16611Documento225 páginasMunsch Washington 0250E 16611taliamatAinda não há avaliações
- Hull Fouling As A Mechanism For Marine IDocumento59 páginasHull Fouling As A Mechanism For Marine IsoaresacpAinda não há avaliações
- Camiguin National High School: Diversity of Mollusks in Tanguines Lagoon, Benoni, CamiguinDocumento11 páginasCamiguin National High School: Diversity of Mollusks in Tanguines Lagoon, Benoni, CamiguinPRECooLAinda não há avaliações
- Ocean-Aquariums and Their Visitor Experiences: An Instrument For Promoting Tourism and Aquatic Wildlife ConservationDocumento8 páginasOcean-Aquariums and Their Visitor Experiences: An Instrument For Promoting Tourism and Aquatic Wildlife ConservationInternational Journal of Innovative Science and Research Technology100% (1)
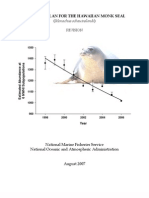
- Hawaiian Monk SealDocumento165 páginasHawaiian Monk SealTrisha Kehaulani Watson-SproatAinda não há avaliações
- Proposal DraftDocumento14 páginasProposal Draftapi-433033817Ainda não há avaliações
- Sphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkDocumento13 páginasSphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkRodrigo MazzoleniAinda não há avaliações
- Contenido Estomacal USA FloridaDocumento19 páginasContenido Estomacal USA FloridaRuth Vasquez LevyAinda não há avaliações
- WIF26 17 SinghDocumento7 páginasWIF26 17 SinghMaybel BalansagAinda não há avaliações
- Monterey Bay Aquarium Member Magazine Shorelines Spring 2011Documento16 páginasMonterey Bay Aquarium Member Magazine Shorelines Spring 2011Monterey Bay AquariumAinda não há avaliações
- Pez Maya Magazine Vol 1Documento16 páginasPez Maya Magazine Vol 1GVI MexicoAinda não há avaliações
- Aquarius User Manual: Florida International University (FIU)Documento11 páginasAquarius User Manual: Florida International University (FIU)Divers Dream PrahaAinda não há avaliações
- Biology and Blooms of The Edible Jellyfish in The Saleh Bay, IndonesiaDocumento14 páginasBiology and Blooms of The Edible Jellyfish in The Saleh Bay, Indonesiabuya301290Ainda não há avaliações
- Kapaa High SchoolDocumento1 páginaKapaa High SchoolHawaii News NowAinda não há avaliações
- NTSB Preliminary Report On Mokulele PlaneDocumento3 páginasNTSB Preliminary Report On Mokulele PlaneHawaii News NowAinda não há avaliações
- 8-6 Big Island IndictmentDocumento8 páginas8-6 Big Island IndictmentHawaii News Now100% (1)
- Name LogDocumento4 páginasName LogNicoleAinda não há avaliações
- What To Know Before You GoDocumento7 páginasWhat To Know Before You GoHawaii News NowAinda não há avaliações
- Kealoha Divorce PaperworkDocumento5 páginasKealoha Divorce PaperworkHawaii News NowAinda não há avaliações
- 12-24 Doctor ShortageDocumento18 páginas12-24 Doctor ShortageHawaii News NowAinda não há avaliações
- National Transportation Safety Board Aviation Accident Preliminary ReportDocumento2 páginasNational Transportation Safety Board Aviation Accident Preliminary ReportSandra Oshiro100% (1)
- SFF List With Cover Letter 051419 - CaseyDocumento9 páginasSFF List With Cover Letter 051419 - CaseyAnonymous WCaY6rAinda não há avaliações
- Report: Number of Homeless People Living On The Streets of Kauai Nearly Doubled Since Last YearDocumento6 páginasReport: Number of Homeless People Living On The Streets of Kauai Nearly Doubled Since Last YearHawaii News NowAinda não há avaliações
- Kealoha Detention MotionDocumento8 páginasKealoha Detention MotionHawaii News Now0% (1)
- Monster X Tour InfoDocumento5 páginasMonster X Tour InfoHawaii News NowAinda não há avaliações
- Workshop & TaxesDocumento1 páginaWorkshop & TaxesHawaii News NowAinda não há avaliações
- Letter From Maui Community Correctional CenterDocumento2 páginasLetter From Maui Community Correctional CenterHawaii News NowAinda não há avaliações
- What You Need To Know Before The Bruno Mars ConcertDocumento5 páginasWhat You Need To Know Before The Bruno Mars ConcertHawaii News Now100% (4)
- Final Ballot Nominees For The 2019 Na Hoku Hanohano AwardsDocumento6 páginasFinal Ballot Nominees For The 2019 Na Hoku Hanohano AwardsHawaii News NowAinda não há avaliações
- Neighborhoods of PearlridgeDocumento1 páginaNeighborhoods of PearlridgeHawaii News NowAinda não há avaliações
- Audit of The Honolulu Rail ProjectDocumento56 páginasAudit of The Honolulu Rail ProjectHawaii News NowAinda não há avaliações
- Ku Mai Ka Hula 2017 WinnersDocumento2 páginasKu Mai Ka Hula 2017 WinnersHawaii News NowAinda não há avaliações
- H-1 Eastbound Piikoi Closure AnalysisDocumento8 páginasH-1 Eastbound Piikoi Closure AnalysisHawaii News NowAinda não há avaliações
- Audit of The Honolulu Rail ProjectDocumento56 páginasAudit of The Honolulu Rail ProjectHawaii News NowAinda não há avaliações
- Louis and Katherine Kealoha IndictmentDocumento42 páginasLouis and Katherine Kealoha IndictmentHonolulu Star-Advertiser100% (2)
- NOAA Status Report On Coral ReefsDocumento7 páginasNOAA Status Report On Coral ReefsHawaii News NowAinda não há avaliações
- Hokulea Visits The North ShoreDocumento3 páginasHokulea Visits The North ShoreHawaii News NowAinda não há avaliações
- Site Plan: Blaisdell Center Master Plan // July 2017Documento2 páginasSite Plan: Blaisdell Center Master Plan // July 2017Hawaii News NowAinda não há avaliações
- Pan-Pacific Festival EventsDocumento1 páginaPan-Pacific Festival EventsHawaii News NowAinda não há avaliações
- Hawai'i Volcanoes National Park - Bird Species List: Common Name Scientific Name Residency Abundance NotesDocumento4 páginasHawai'i Volcanoes National Park - Bird Species List: Common Name Scientific Name Residency Abundance NotesHawaii News NowAinda não há avaliações
- Hawai'i Volcanoes National Park - Bird Species List: Common Name Scientific Name Residency Abundance NotesDocumento4 páginasHawai'i Volcanoes National Park - Bird Species List: Common Name Scientific Name Residency Abundance NotesHawaii News NowAinda não há avaliações
- 9th Circuit Opinion Upholding Travel Order Injunction - June 12 2017Documento86 páginas9th Circuit Opinion Upholding Travel Order Injunction - June 12 2017Legal InsurrectionAinda não há avaliações
- Full NTSB Report For Makaha Plane CrashDocumento6 páginasFull NTSB Report For Makaha Plane CrashHawaii News NowAinda não há avaliações
- Atoma Amd Mol&Us CCTK) : 2Nd ErmDocumento4 páginasAtoma Amd Mol&Us CCTK) : 2Nd ErmjanviAinda não há avaliações
- Jurnal Job DescriptionDocumento13 páginasJurnal Job DescriptionAji Mulia PrasAinda não há avaliações
- ISO Position ToleranceDocumento15 páginasISO Position ToleranceНиколай КалугинAinda não há avaliações
- How To Install Metal LathDocumento2 páginasHow To Install Metal LathKfir BenishtiAinda não há avaliações
- There Will Come Soft RainsDocumento8 páginasThere Will Come Soft RainsEng ProfAinda não há avaliações
- Thesis TipsDocumento57 páginasThesis TipsJohn Roldan BuhayAinda não há avaliações
- Pam8610 PDFDocumento15 páginasPam8610 PDFRaka Satria PradanaAinda não há avaliações
- Stress Management HandoutsDocumento3 páginasStress Management HandoutsUsha SharmaAinda não há avaliações
- Quality Standards For ECCE INDIA PDFDocumento41 páginasQuality Standards For ECCE INDIA PDFMaryam Ben100% (4)
- Unit 1 PrinciplesDocumento17 páginasUnit 1 PrinciplesRohit YadavAinda não há avaliações
- Governance Operating Model: Structure Oversight Responsibilities Talent and Culture Infrastructu REDocumento6 páginasGovernance Operating Model: Structure Oversight Responsibilities Talent and Culture Infrastructu REBob SolísAinda não há avaliações
- Department of Ece, Adhiparasakthi College of Engineering, KalavaiDocumento31 páginasDepartment of Ece, Adhiparasakthi College of Engineering, KalavaiGiri PrasadAinda não há avaliações
- Load Chart Crane LiftingDocumento25 páginasLoad Chart Crane LiftingLauren'sclub EnglishBimbel Sd-sma100% (1)
- Industrial ReportDocumento52 páginasIndustrial ReportSiddharthAinda não há avaliações
- 28 TerminalsDocumento42 páginas28 TerminalsAlin FazacasAinda não há avaliações
- HRM Practices in NepalDocumento22 páginasHRM Practices in NepalBodhiAinda não há avaliações
- JO 20221109 NationalDocumento244 páginasJO 20221109 NationalMark Leo BejeminoAinda não há avaliações
- 2.0. Design, Protection and Sizing of Low Voltage Electrical Installations (Wiring) To IEE Wiring Regulations BS 7671 MS IEC (IEC) 60364 - 16&17.12.15Documento2 páginas2.0. Design, Protection and Sizing of Low Voltage Electrical Installations (Wiring) To IEE Wiring Regulations BS 7671 MS IEC (IEC) 60364 - 16&17.12.15Edison LimAinda não há avaliações
- FT2020Documento7 páginasFT2020Sam SparksAinda não há avaliações
- CSWIP-WP-19-08 Review of Welding Procedures 2nd Edition February 2017Documento6 páginasCSWIP-WP-19-08 Review of Welding Procedures 2nd Edition February 2017oberai100% (1)
- WWW Ranker Com List Best-Isekai-Manga-Recommendations Ranker-AnimeDocumento8 páginasWWW Ranker Com List Best-Isekai-Manga-Recommendations Ranker-AnimeDestiny EasonAinda não há avaliações
- Moral GPS: A Positive Ethic For Our Multicultural WorldDocumento279 páginasMoral GPS: A Positive Ethic For Our Multicultural WorldLeonard Bowman75% (4)
- My BaboogDocumento1 páginaMy BaboogMaral Habeshian VieiraAinda não há avaliações
- Mahindra&mahindraDocumento95 páginasMahindra&mahindraAshik R GowdaAinda não há avaliações
- Etag 002 PT 2 PDFDocumento13 páginasEtag 002 PT 2 PDFRui RibeiroAinda não há avaliações
- Engineering Management: Class RequirementsDocumento30 páginasEngineering Management: Class RequirementsMigaeaAinda não há avaliações
- 123Documento3 páginas123Phoebe AradoAinda não há avaliações
- Management PriniciplesDocumento87 páginasManagement Priniciplesbusyboy_spAinda não há avaliações
- P3 Past Papers Model AnswersDocumento211 páginasP3 Past Papers Model AnswersEyad UsamaAinda não há avaliações
- CIPD L5 EML LOL Wk3 v1.1Documento19 páginasCIPD L5 EML LOL Wk3 v1.1JulianAinda não há avaliações
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessAinda não há avaliações
- The Other End of the Leash: Why We Do What We Do Around DogsNo EverandThe Other End of the Leash: Why We Do What We Do Around DogsNota: 5 de 5 estrelas5/5 (65)
- Will's Red Coat: The Story of One Old Dog Who Chose to Live AgainNo EverandWill's Red Coat: The Story of One Old Dog Who Chose to Live AgainNota: 4.5 de 5 estrelas4.5/5 (18)
- The Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationNo EverandThe Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationNota: 4 de 5 estrelas4/5 (37)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNo EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNota: 5 de 5 estrelas5/5 (3)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNo EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNota: 4.5 de 5 estrelas4.5/5 (24)
- Come Back, Como: Winning the Heart of a Reluctant DogNo EverandCome Back, Como: Winning the Heart of a Reluctant DogNota: 3.5 de 5 estrelas3.5/5 (10)
- Inside of a Dog: What Dogs See, Smell, and KnowNo EverandInside of a Dog: What Dogs See, Smell, and KnowNota: 4 de 5 estrelas4/5 (390)
- An Eagle Named Freedom: My True Story of a Remarkable FriendshipNo EverandAn Eagle Named Freedom: My True Story of a Remarkable FriendshipAinda não há avaliações
- Show Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredNo EverandShow Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredNota: 3.5 de 5 estrelas3.5/5 (13)
- Dogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowNo EverandDogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowAinda não há avaliações
- The Wrong Dog: An Unlikely Tale of Unconditional LoveNo EverandThe Wrong Dog: An Unlikely Tale of Unconditional LoveNota: 4.5 de 5 estrelas4.5/5 (26)
- Second Chances: A Marine, His Dog, and Finding RedemptionNo EverandSecond Chances: A Marine, His Dog, and Finding RedemptionAinda não há avaliações
- MINE!: A PRACTICAL GUIDE TO RESOURCE GUARDING IN DOGSNo EverandMINE!: A PRACTICAL GUIDE TO RESOURCE GUARDING IN DOGSNota: 4 de 5 estrelas4/5 (15)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsAinda não há avaliações
- Puppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!No EverandPuppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!Nota: 5 de 5 estrelas5/5 (85)
- Arthur: The Dog Who Crossed the Jungle to Find a HomeNo EverandArthur: The Dog Who Crossed the Jungle to Find a HomeNota: 4.5 de 5 estrelas4.5/5 (20)
- Edward's Menagerie: Dogs: 50 canine crochet patternsNo EverandEdward's Menagerie: Dogs: 50 canine crochet patternsNota: 3 de 5 estrelas3/5 (5)
- Your Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesNo EverandYour Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesNota: 4 de 5 estrelas4/5 (31)
- Tamed and Untamed: Close Encounters of the Animal KindNo EverandTamed and Untamed: Close Encounters of the Animal KindNota: 3.5 de 5 estrelas3.5/5 (32)
- The Good Good Pig: The Extraordinary Life of Christopher HogwoodNo EverandThe Good Good Pig: The Extraordinary Life of Christopher HogwoodNota: 4 de 5 estrelas4/5 (282)
- The Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenNo EverandThe Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenNota: 4.5 de 5 estrelas4.5/5 (49)
- Meet Your Dog: The Game-Changing Guide to Understanding Your Dog's BehaviorNo EverandMeet Your Dog: The Game-Changing Guide to Understanding Your Dog's BehaviorNota: 5 de 5 estrelas5/5 (34)