Você também pode gostar
- Reacción de ArthusDocumento6 páginasReacción de ArthusGloria María100% (2)
- Reparación de Tejidos - RobbinsDocumento14 páginasReparación de Tejidos - RobbinsGerson OrellanaAinda não há avaliações
- Tipos de BiopsiasDocumento2 páginasTipos de BiopsiasÁngel García Hernández80% (5)
- Equipo 8 Factor Estimulante de Colonias Ftp02a 20-2Documento9 páginasEquipo 8 Factor Estimulante de Colonias Ftp02a 20-2Ruth Cobo RosalesAinda não há avaliações
- Resumen Neoplasias Linfoides y Mieloides. Patología RobbinsDocumento39 páginasResumen Neoplasias Linfoides y Mieloides. Patología RobbinsAilyn ForeroAinda não há avaliações
- Reparación de TejidosDocumento8 páginasReparación de TejidosFernando VillavicencioAinda não há avaliações
- Anemia FanconiDocumento12 páginasAnemia FanconiDayana Cañas FlorezAinda não há avaliações
- LinfopoyesisDocumento12 páginasLinfopoyesisViianey Navii SpoockAinda não há avaliações
- Fisiopatologia Leucemia Mieloide AgudaDocumento2 páginasFisiopatologia Leucemia Mieloide AgudaShanel Marithain100% (2)
- Practica de FagocitosisDocumento2 páginasPractica de FagocitosisCelitzy GómezAinda não há avaliações
- Trastornos Hemostasia Primaria y Secundaria. AlgoritmoDocumento2 páginasTrastornos Hemostasia Primaria y Secundaria. AlgoritmoAlex PazAinda não há avaliações
- Interpretación Examen General de OrinaDocumento67 páginasInterpretación Examen General de OrinaJulia RuizAinda não há avaliações
- Metodos de Estudio de La PatologiaDocumento8 páginasMetodos de Estudio de La PatologiaTeresa Montes33% (3)
- Métodos de Estudio de La Anatomía PatológicaDocumento3 páginasMétodos de Estudio de La Anatomía PatológicaWendyGianella100% (1)
- Linfoma Folicular 1Documento11 páginasLinfoma Folicular 1SHEYLA SHARINA FUENTES PINTOAinda não há avaliações
- Generalidades y Semiología Del Niño Sano EQUIPO 1Documento110 páginasGeneralidades y Semiología Del Niño Sano EQUIPO 111 Francisco Fierro Olea100% (1)
- Linfoma No HodgkinDocumento44 páginasLinfoma No HodgkinMENDOZA REYES LALY TATIANA100% (1)
- Interrogatorio Cabeza y Cuello - Órganos SensorialesDocumento34 páginasInterrogatorio Cabeza y Cuello - Órganos SensorialesMiriamFuentesAinda não há avaliações
- Liquido PeritonealDocumento12 páginasLiquido PeritonealQUIMICO CLINICO WILLIANS SANCHEZ100% (10)
- InterleucinasDocumento2 páginasInterleucinasJosé Damián67% (3)
- Fisiopatologia de Evc HemorragicoDocumento18 páginasFisiopatologia de Evc HemorragicovalfreAinda não há avaliações
- Sistema Bethesda en Citopatologia de TiroidesDocumento41 páginasSistema Bethesda en Citopatologia de TiroidesJuanJo Borja100% (1)
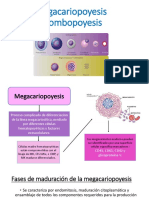
- Megacariopoyesis y TrombopoyesisDocumento5 páginasMegacariopoyesis y TrombopoyesisKatherine Chavez67% (3)
- Tema 6 Enfermedades de Los LeucocitosDocumento73 páginasTema 6 Enfermedades de Los LeucocitosHJ GuimasAinda não há avaliações
- Anemias Hemolíticas Extracorpusculares o ExtrínsecasDocumento7 páginasAnemias Hemolíticas Extracorpusculares o ExtrínsecasAndreaRosales0% (1)
- Síndrome de Absorción Intestinal DeficienteDocumento19 páginasSíndrome de Absorción Intestinal DeficienteVania100% (1)
- Inmunohistoquímica Del HígadoDocumento12 páginasInmunohistoquímica Del HígadoAndrea GuanoluisaAinda não há avaliações
- Infeccion Por ChancroideDocumento24 páginasInfeccion Por ChancroideClau TomAinda não há avaliações
- Eritrocitos y HemoglobinaDocumento20 páginasEritrocitos y HemoglobinaJulio CruzAinda não há avaliações
- Citodiagnóstico ClínicoDocumento19 páginasCitodiagnóstico ClínicoMaria Concepción Chura BarónAinda não há avaliações
- HematopoyesisDocumento32 páginasHematopoyesisbeto vil100% (2)
- LLA FinalDocumento19 páginasLLA FinalOrnellaAinda não há avaliações
- MielopoyesisDocumento9 páginasMielopoyesisLeidy GonzabayAinda não há avaliações
- Terapéutica EmpleadaDocumento19 páginasTerapéutica EmpleadaAngelica Cg100% (2)
- Aislamiento de Células Mononucleares Humanas Por Gradiente de FicollDocumento3 páginasAislamiento de Células Mononucleares Humanas Por Gradiente de FicollJoséLuisAguirreAinda não há avaliações
- Examen general de orina completoDocumento72 páginasExamen general de orina completohermiz bolaños100% (1)
- BASÓFILOSDocumento9 páginasBASÓFILOSIsrael Vargas LeonAinda não há avaliações
- Biometría HemáticaDocumento24 páginasBiometría HemáticaMontseCandiaTeapilaAinda não há avaliações
- Hematología Biometria Hematica Estudio Resultado Valores de ReferenciaDocumento3 páginasHematología Biometria Hematica Estudio Resultado Valores de ReferenciaYeseniaa DuarteeAinda não há avaliações
- NEOPLASIASDocumento17 páginasNEOPLASIASManuel Magaña100% (1)
- Citología de OrinaDocumento53 páginasCitología de OrinaFrancisca50% (2)
- Capitulo 17 - Tubo DigestivoDocumento7 páginasCapitulo 17 - Tubo DigestivoJuli Ferreyra Marchuk83% (12)
- Vias LagrimalesDocumento9 páginasVias LagrimalesREYES AGUILAR SALINASAinda não há avaliações
- Fisiopatología - Porth - 7ed PDFDocumento1.568 páginasFisiopatología - Porth - 7ed PDFJr Adrian Bedregal Gomez90% (10)
- Atlas de Hematología Clínica: Portada AutoresDocumento2 páginasAtlas de Hematología Clínica: Portada AutoresAlan NavaAinda não há avaliações
- Sindrome HemorragiparoDocumento50 páginasSindrome HemorragiparoMarco MoralesAinda não há avaliações
- Cuestionario Patologia de RobbinsDocumento4 páginasCuestionario Patologia de RobbinsIsa FabaraAinda não há avaliações
- Cicatrizacion y Reparacion - Robbins - ResumenDocumento19 páginasCicatrizacion y Reparacion - Robbins - ResumenPaula Andrea Garcés Quitumbo100% (1)
- Sabiston CicatrizaciónDocumento33 páginasSabiston CicatrizaciónEdwin Lionel Castillo MirandaAinda não há avaliações
- Microambiente de La Medula OseaDocumento9 páginasMicroambiente de La Medula OseaChristian Correa Verde100% (1)
- Enfermedad de Still Del Adulto (ESA)Documento14 páginasEnfermedad de Still Del Adulto (ESA)José Fernando Huanca CordovaAinda não há avaliações
- MielopoyesisDocumento4 páginasMielopoyesisMariangel Jimenez100% (1)
- Hipermutacion SomaticaDocumento2 páginasHipermutacion SomaticaIvan Moya50% (2)
- Formación de plaquetas sanguíneasDocumento5 páginasFormación de plaquetas sanguíneasAlice0% (1)
- HematopoyesisDocumento19 páginasHematopoyesisJocelyne Calderon PadillaAinda não há avaliações
- Hematopoyesis MonseDocumento5 páginasHematopoyesis Monsejuan Cordova LaraAinda não há avaliações
- Hematopoyesis ResumenDocumento4 páginasHematopoyesis ResumenErika Elizabeth Medina ToledoAinda não há avaliações
- HEMATOPOYESISDocumento7 páginasHEMATOPOYESISAileen PresuelAinda não há avaliações
- Articulo 2 HematologiaDocumento10 páginasArticulo 2 HematologiaRene Gtz SAinda não há avaliações
- Celulas MadreDocumento9 páginasCelulas Madrearmanditocote94Ainda não há avaliações
- Revista Conocimiento 21Documento76 páginasRevista Conocimiento 21Revista Conocimiento en línea.100% (1)
- E1 Papel de Los Ietabolitos Secundarios en La Utilizadcei ÓnDocumento77 páginasE1 Papel de Los Ietabolitos Secundarios en La Utilizadcei ÓnMarlene Sinai MeloAinda não há avaliações
- 1 Editorial Muerte CerebralDocumento3 páginas1 Editorial Muerte CerebralMarlene Sinai MeloAinda não há avaliações
- 9 PapeldelafuncionDocumento8 páginas9 PapeldelafuncionMarlene Sinai MeloAinda não há avaliações
- Biotransformacion FarmacosDocumento24 páginasBiotransformacion FarmacosMarlene Sinai MeloAinda não há avaliações
- TRabajo LidocainaDocumento7 páginasTRabajo LidocainaMarlene Sinai MeloAinda não há avaliações
- Análisis Químico de Plantas MedicinalesDocumento13 páginasAnálisis Químico de Plantas MedicinalesMarlene Sinai MeloAinda não há avaliações
- Cuadro ABCDocumento2 páginasCuadro ABCMarlene Sinai MeloAinda não há avaliações
- Preguntas AnamnesisDocumento12 páginasPreguntas AnamnesisSusana AlvaradoAinda não há avaliações
- Evaluacion Del Educando Con Dificultades Del Aprendizaje. Cuadros Comparativos de ConceptosDocumento7 páginasEvaluacion Del Educando Con Dificultades Del Aprendizaje. Cuadros Comparativos de ConceptossanysAinda não há avaliações
- Enfermedades y BacteriasDocumento50 páginasEnfermedades y BacteriasosoposoloAinda não há avaliações
- Atención A Las Nee en El Aula Regular Romero & BritoDocumento25 páginasAtención A Las Nee en El Aula Regular Romero & BritoJosé Ávila BAinda não há avaliações
- Faramacos Supresores LactanciaDocumento18 páginasFaramacos Supresores LactanciaJUANA100% (1)
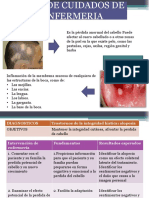
- EnfermeriaDocumento11 páginasEnfermeriaAda Cuquian Borda100% (1)
- Colegio Comfandi: Síntesis de proteínas hemoglobinaDocumento5 páginasColegio Comfandi: Síntesis de proteínas hemoglobinaSandra Patricia Carreño GutierrezAinda não há avaliações
- Informe # 11 Meiosis - HerenciaDocumento3 páginasInforme # 11 Meiosis - HerenciaMishellAlvarez100% (1)
- Cual Es La Accion de La Solucion Fijadora Sobre Estructuras CelularesDocumento2 páginasCual Es La Accion de La Solucion Fijadora Sobre Estructuras CelularesLuisaArismendyAinda não há avaliações
- AlcaptonuriaDocumento12 páginasAlcaptonuriaasfbisAinda não há avaliações
- La Maté Porque Era Mía Psicobiología de La Ira, De... - (PG 78 - 91)Documento14 páginasLa Maté Porque Era Mía Psicobiología de La Ira, De... - (PG 78 - 91)leidy lorena angel gomez100% (2)
- Admisión - 6°Documento16 páginasAdmisión - 6°martha florezAinda não há avaliações
- Páginas Web TOCDocumento9 páginasPáginas Web TOCdidi monroeAinda não há avaliações
- 678 Monjas y Un CientíficoDocumento1 página678 Monjas y Un CientíficoAnibal Fernandez OliveiraAinda não há avaliações
- El clítoris, el secreto del placer femeninoDocumento5 páginasEl clítoris, el secreto del placer femeninofrankx_Ainda não há avaliações
- Marco Teorico SatisfaccionDocumento6 páginasMarco Teorico SatisfaccionMichael Aquino Herrera100% (1)
- PREGUNTASDocumento2 páginasPREGUNTASAnthony SanchezAinda não há avaliações
- Protocolo bacterias oportunistasDocumento8 páginasProtocolo bacterias oportunistasNiki VRAinda não há avaliações
- Aislamiento de Caseína de LecheDocumento3 páginasAislamiento de Caseína de LecheAlejandra De HernandezAinda não há avaliações
- Distintas Plagas y Enfermedades en Las PlantasDocumento5 páginasDistintas Plagas y Enfermedades en Las PlantasMoises Sojo SaavedraAinda não há avaliações
- Factores de riesgo y teorías sobre el suicidioDocumento2 páginasFactores de riesgo y teorías sobre el suicidioAngelaa G Oviedo Carquin80% (5)
- Tendinitis Rotuliana: Etiología, Cuadro Clínico, Tratamiento y PrevenciónDocumento3 páginasTendinitis Rotuliana: Etiología, Cuadro Clínico, Tratamiento y PrevenciónUri Claro100% (1)
- Fibrosis PulmonarDocumento24 páginasFibrosis PulmonarAmir Llontop Sernaque100% (1)
- La Armonía en Los Dedos de La ManoDocumento4 páginasLa Armonía en Los Dedos de La ManomaguiaraveAinda não há avaliações
- Efecto de La Temperatura Sobre Las Membranas BiologicasDocumento17 páginasEfecto de La Temperatura Sobre Las Membranas BiologicasWilmer Javier100% (1)
- Fisiología de La Diferenciación SexualDocumento16 páginasFisiología de La Diferenciación SexualSamuel Fernando Velasco Chacon0% (1)
- Lectura Emocional DR Franco RossomandoDocumento126 páginasLectura Emocional DR Franco RossomandoBilance100% (2)
- Normas de EnfermeríaDocumento3 páginasNormas de EnfermeríaMickel Novas100% (2)
- Adán Cromosomal YDocumento4 páginasAdán Cromosomal Yarturoalcides8171100% (1)
- Células Troncales Mesenquimales, Revisión Bibliografica, RCG, Grupo Central Regenerativo, CBCellsDocumento18 páginasCélulas Troncales Mesenquimales, Revisión Bibliografica, RCG, Grupo Central Regenerativo, CBCellsangel 1958Ainda não há avaliações