Você também pode gostar
- Dimensiones de la exclusión psicosocial.: Elementos para la teoría, la investigación y la intervenciónNo EverandDimensiones de la exclusión psicosocial.: Elementos para la teoría, la investigación y la intervenciónAinda não há avaliações
- Mapa MentalDocumento2 páginasMapa MentalNaty Bernaola MelendezAinda não há avaliações
- AnemiaDocumento29 páginasAnemiaMarilu VargasAinda não há avaliações
- Ensayo El Cerebro Social.Documento6 páginasEnsayo El Cerebro Social.Derly Yohanna CUEVAS GALLOAinda não há avaliações
- 6-Sandro-y-Mitzy - TABULACIONESDocumento57 páginas6-Sandro-y-Mitzy - TABULACIONESCarlos Arturo Rueda RamirezAinda não há avaliações
- Tania Ayala Silvia Pajuelo Tesis Titulo Profesional 2018Documento205 páginasTania Ayala Silvia Pajuelo Tesis Titulo Profesional 2018Licha CarMonAinda não há avaliações
- Tesis Sentido de Comunidad y Bienestar PsicologicoDocumento143 páginasTesis Sentido de Comunidad y Bienestar PsicologicoKay MirandaAinda não há avaliações
- Charlas de 5 Minutos PDFDocumento32 páginasCharlas de 5 Minutos PDFAngiolo Puentes NievesAinda não há avaliações
- Ensayo Sobre La Hipótesis Del Cerebro SocialDocumento5 páginasEnsayo Sobre La Hipótesis Del Cerebro SocialMaira Alejandra EspitiaAinda não há avaliações
- Funcionamiento Familiar InvestigacionDocumento44 páginasFuncionamiento Familiar InvestigacionAngeluz FiorellaAinda não há avaliações
- Charla de Seguridad de 5 Minutos 8Documento1 páginaCharla de Seguridad de 5 Minutos 8leyisap02100% (1)
- ATRIBUCIÓN Diapositivas PDFDocumento12 páginasATRIBUCIÓN Diapositivas PDFMiguel Ángel TórrezAinda não há avaliações
- Monografia de PSICOLOGÍA POSITIVADocumento20 páginasMonografia de PSICOLOGÍA POSITIVAElio Lazo ChayanAinda não há avaliações
- Plan de TrabajoDocumento9 páginasPlan de TrabajoGaar NewAinda não há avaliações
- Psicología AmbientalDocumento16 páginasPsicología Ambientalpaula maria ortiz100% (2)
- La Importancia Del Juego en La FamiliaDocumento2 páginasLa Importancia Del Juego en La FamiliaJuan CarlosAinda não há avaliações
- Psicologia Comunitaria y AmbientalDocumento24 páginasPsicologia Comunitaria y AmbientalIndira MojicaAinda não há avaliações
- Psicologia PositivaDocumento16 páginasPsicologia PositivaYesenia Fuentes ValdiviaAinda não há avaliações
- Resumen Cerebro SocialDocumento6 páginasResumen Cerebro SocialLauraAinda não há avaliações
- Estres en NeonatosDocumento10 páginasEstres en Neonatossolmassi100% (1)
- Proyecto de Intervencion OrganizacionalDocumento10 páginasProyecto de Intervencion Organizacionalmayo ketchupAinda não há avaliações
- Resumen Final Psico. GeneralDocumento87 páginasResumen Final Psico. Generaljudith coronelAinda não há avaliações
- Matriz de Habilidades Socioemocionales ClaveDocumento1 páginaMatriz de Habilidades Socioemocionales ClavevallocoAinda não há avaliações
- Técnicas de Manejo de Conflictos y Análisis de NegociaciónDocumento2 páginasTécnicas de Manejo de Conflictos y Análisis de Negociación10625095Ainda não há avaliações
- Funcionamiento FamiliarDocumento3 páginasFuncionamiento FamiliarJhoana PiconAinda não há avaliações
- Resumen de Psicología EducativaDocumento3 páginasResumen de Psicología Educativaroxana ucedoAinda não há avaliações
- Psicologia AmbientalDocumento22 páginasPsicologia Ambientalmil_ric145657Ainda não há avaliações
- Competencias Socioemocionale - Pensamiento Emocion y AccionDocumento4 páginasCompetencias Socioemocionale - Pensamiento Emocion y AccionNADIA SONIA CARPIO CORTEZAinda não há avaliações
- Desarrollar La Autonomía PersonalDocumento2 páginasDesarrollar La Autonomía PersonalSaÄmy GõmëzAinda não há avaliações
- Monografia Psicologia AmbientalDocumento16 páginasMonografia Psicologia AmbientalyefriblueAinda não há avaliações
- 8 Habilidades SocialesDocumento35 páginas8 Habilidades SocialesGaby Ibarrola MurilloAinda não há avaliações
- Tarea 2 T. FamiliarDocumento10 páginasTarea 2 T. FamiliarRramonita lamiz100% (1)
- Trabajo de Practicas Profesionales InpecDocumento36 páginasTrabajo de Practicas Profesionales Inpecluis lopezAinda não há avaliações
- Programa de Intervención para El Desarrollo (Caso 2)Documento21 páginasPrograma de Intervención para El Desarrollo (Caso 2)Gabriel ApazaAinda não há avaliações
- PAT Monguete 2019Documento19 páginasPAT Monguete 2019Haydee D L CruzAinda não há avaliações
- Importancia de La PsicologiaDocumento7 páginasImportancia de La PsicologiaCesar QuicazanAinda não há avaliações
- Diagnóstico Situacional - TutoriaDocumento1 páginaDiagnóstico Situacional - TutoriaKatherine Ossandon VizquerraAinda não há avaliações
- Prevención de Las Alteraciones Psicológicas en Niños y AdolescentesDocumento4 páginasPrevención de Las Alteraciones Psicológicas en Niños y AdolescentesJhon Lauren'sAinda não há avaliações
- Modulo de Consejeria PsicologicaDocumento61 páginasModulo de Consejeria PsicologicaRenzo GuillermoAinda não há avaliações
- Ansiedad en Poblaciones de Estudiantes Universitarios Argentinos y Estadounidense. Recolección Muestral Via Internet y PresencialDocumento199 páginasAnsiedad en Poblaciones de Estudiantes Universitarios Argentinos y Estadounidense. Recolección Muestral Via Internet y PresencialKeny IllanesAinda não há avaliações
- Beneficios de La Educacion FisicaDocumento17 páginasBeneficios de La Educacion Fisicatiby79Ainda não há avaliações
- Neuropsicologia - Tarea - 1 - Pre-Esaberes - Dianabarrios.Documento10 páginasNeuropsicologia - Tarea - 1 - Pre-Esaberes - Dianabarrios.diana barriosAinda não há avaliações
- Investigación A Un PsicólogoDocumento14 páginasInvestigación A Un PsicólogoFernando Villaseca CorreaAinda não há avaliações
- Practica 5f1Documento27 páginasPractica 5f1Jack SkellingtonAinda não há avaliações
- NeuropsicologiaDocumento3 páginasNeuropsicologiaAngie Bravo100% (1)
- Psicologia OrganizacionalDocumento18 páginasPsicologia OrganizacionalAllyson Delgado QuispeAinda não há avaliações
- Proyecto Cuantitativa Apego PDFDocumento41 páginasProyecto Cuantitativa Apego PDFSthefany Bautista100% (1)
- Ps Organizacional SEPARATA Prevención y PromociónDocumento25 páginasPs Organizacional SEPARATA Prevención y PromociónCarlos Andrés NeyraAinda não há avaliações
- La Mente SabiaDocumento5 páginasLa Mente SabiaCristian Ayala RománAinda não há avaliações
- Inteligencia EmocionalDocumento25 páginasInteligencia EmocionalLuzma GarciaAinda não há avaliações
- Psicologia de Los Grupos Act 3Documento13 páginasPsicologia de Los Grupos Act 3LUZMALAS100% (1)
- Psicologia ComunitariaDocumento17 páginasPsicologia ComunitariaSheyla VelozAinda não há avaliações
- Plan de Accion TutoriaDocumento30 páginasPlan de Accion TutoriaJhonatan MatamorosAinda não há avaliações
- EnsayoDocumento9 páginasEnsayoJuan Ramirez DominguezAinda não há avaliações
- Luis Emiro SotoDocumento18 páginasLuis Emiro SotoInglys MayoAinda não há avaliações
- Convivencia Familiar PDFDocumento16 páginasConvivencia Familiar PDFELIZABETH CRISTINA GIRALDO MONTOYAAinda não há avaliações
- Autoestima 2Documento14 páginasAutoestima 2Abel Felipe Barrena GurbillnAinda não há avaliações
- Apuntes de Escuela de Milán SintesisDocumento9 páginasApuntes de Escuela de Milán SintesisBig DiuxAinda não há avaliações
- Ley General de Las Personas Con DiscapacidadDocumento17 páginasLey General de Las Personas Con DiscapacidadMyle AbrocAinda não há avaliações
- Acedo&gomila DunbarDocumento22 páginasAcedo&gomila DunbarToni GomilaAinda não há avaliações
- ¿Adicciones... Sin Drogas. Las Nuevas AdiccionesDocumento125 páginas¿Adicciones... Sin Drogas. Las Nuevas AdiccionesHector Garduño Garduño100% (1)
- Atencion y Trat en Prision Por Uso Drogas 2012Documento290 páginasAtencion y Trat en Prision Por Uso Drogas 2012alffix100% (1)
- El Cambio en Las Conductas AdictivasDocumento14 páginasEl Cambio en Las Conductas AdictivasMarjorie LunaAinda não há avaliações
- Adicciones Conocimiento, Atención Integrada y Acción Preventiva - Iñaki Marke PDFDocumento561 páginasAdicciones Conocimiento, Atención Integrada y Acción Preventiva - Iñaki Marke PDFGabriel CabreraAinda não há avaliações
- Manual Del Test de Colores de LuscherDocumento108 páginasManual Del Test de Colores de LuscherRuth Obregon92% (181)
- Alcohol y Drogas Prevención en Adolescentes - MÉdico Francisco A. Cantú PDFDocumento65 páginasAlcohol y Drogas Prevención en Adolescentes - MÉdico Francisco A. Cantú PDFjesus ruizAinda não há avaliações
- Pichon Riviere Teoria Del VinculoDocumento64 páginasPichon Riviere Teoria Del VinculoyecowAinda não há avaliações
- BowenDocumento22 páginasBowenPriscila Heredia Ramos100% (1)
- 5000 Frases Precocinadas para Textos Científicos PDFDocumento116 páginas5000 Frases Precocinadas para Textos Científicos PDFEmmanuelCastilloAinda não há avaliações
- Mentalizacion de Los AfectosDocumento16 páginasMentalizacion de Los AfectosparparcitaAinda não há avaliações
- AutomutilacionDocumento7 páginasAutomutilacionVanne OlivaAinda não há avaliações
- Meditaciones MetafísicasDocumento53 páginasMeditaciones Metafísicaspablotherabbit90% (10)
- Conferencia 17 El Sentido de Los Sintomas FreudDocumento8 páginasConferencia 17 El Sentido de Los Sintomas FreudPriscila Heredia RamosAinda não há avaliações
- El Niño y El SignificanteDocumento11 páginasEl Niño y El SignificantePriscila Heredia RamosAinda não há avaliações
- La Expresion de Las Emociones en DarwinDocumento11 páginasLa Expresion de Las Emociones en Darwinapi-370027860% (5)
- Contribuciones Al Debate Sobre El Concepto de Historia RecienteDocumento14 páginasContribuciones Al Debate Sobre El Concepto de Historia RecientePriscila Heredia RamosAinda não há avaliações
- La Expresion de Las Emociones en DarwinDocumento11 páginasLa Expresion de Las Emociones en Darwinapi-370027860% (5)
- Politica Poder y Espacio Público El Pensamiento de Hannan ArendtDocumento12 páginasPolitica Poder y Espacio Público El Pensamiento de Hannan ArendtPriscila Heredia RamosAinda não há avaliações
- Cerebro, Izquierda y DerechaDocumento1 páginaCerebro, Izquierda y DerechaPriscila Heredia RamosAinda não há avaliações
- 85119330-Las Reglas Del Método SociológicoDocumento261 páginas85119330-Las Reglas Del Método SociológicoPriscila Heredia RamosAinda não há avaliações
- Mendoza Publ 2011 Proyecto-Alfa-tuningDocumento9 páginasMendoza Publ 2011 Proyecto-Alfa-tuningOscarCorzoAinda não há avaliações
- BotanicaDocumento8 páginasBotanicaJuan Pablo RMAinda não há avaliações
- Teorías Sobre El Origen Del UniversoDocumento8 páginasTeorías Sobre El Origen Del UniversoJosué CornejoAinda não há avaliações
- Coordenadas Ecuatoriales y LocalesDocumento9 páginasCoordenadas Ecuatoriales y LocalesPablo PillotAinda não há avaliações
- La Malaria en GuatemalaDocumento9 páginasLa Malaria en GuatemalaClaudia MtzAinda não há avaliações
- Examen Vi Nivel Geoquimica DiplomadoDocumento2 páginasExamen Vi Nivel Geoquimica DiplomadoAnonymous JKmMlHJtAinda não há avaliações
- Períodos BotánicosDocumento1 páginaPeríodos BotánicosC AndresAinda não há avaliações
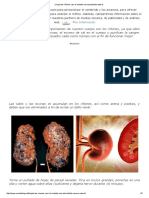
- Limpia Tus Riñones Casi Al Instante Con Esta Bebida NaturalDocumento3 páginasLimpia Tus Riñones Casi Al Instante Con Esta Bebida NaturalOscar Fernandez SanchezAinda não há avaliações
- Lectura - Descubrimiento de La Estructura Del ADNDocumento5 páginasLectura - Descubrimiento de La Estructura Del ADNMarioBrocchiAinda não há avaliações
- Adolescencia y NoviazgoDocumento1 páginaAdolescencia y NoviazgoJorge Luis PerezAinda não há avaliações
- SesquiditerpenosDocumento23 páginasSesquiditerpenosProxy100% (3)
- Contrato Concesion DrummondDocumento3 páginasContrato Concesion DrummondSantiago alvarez ceronAinda não há avaliações
- Ejemplo Secuencia Didactica Ciencias NaturalesDocumento11 páginasEjemplo Secuencia Didactica Ciencias NaturalesYanina Hernandez100% (3)
- Perfil AvesDocumento12 páginasPerfil AvesMilton Apaza PaxiAinda não há avaliações
- Manual Sobre El Manejo de Peligros Naturales en La Planificación para El Desarrollo Regional IntegradoDocumento487 páginasManual Sobre El Manejo de Peligros Naturales en La Planificación para El Desarrollo Regional IntegradoaleydadeysAinda não há avaliações
- White Labs CatalogoEspanol PDFDocumento28 páginasWhite Labs CatalogoEspanol PDFFrancisco LinaresAinda não há avaliações
- Programacion en AutolispDocumento21 páginasProgramacion en AutolispSixto PlasenciaAinda não há avaliações
- Pau Biologia LipidosDocumento5 páginasPau Biologia LipidosmikelemanAinda não há avaliações
- Observaciones Sobre La Entomofauna Asociada A Cinco Cultivares de Amaranto (Amaranthus SPP) CultivadosDocumento46 páginasObservaciones Sobre La Entomofauna Asociada A Cinco Cultivares de Amaranto (Amaranthus SPP) CultivadosMartinAinda não há avaliações
- Practica Trabajo y EnergíaDocumento9 páginasPractica Trabajo y Energíaeliceogimenez100% (3)
- Teoria de La EvolucionDocumento16 páginasTeoria de La EvolucionGeraldin LozanoAinda não há avaliações
- Curriculum Vitae by Al NuevoDocumento3 páginasCurriculum Vitae by Al NuevoVictor Raul Solis GaravitoAinda não há avaliações
- Control EtologicoDocumento7 páginasControl EtologicoMariaElenaMorenoAinda não há avaliações
- Microorganismos Productores de AlcoholDocumento20 páginasMicroorganismos Productores de AlcoholManu San Clemente100% (2)
- AVES DiapositivasDocumento22 páginasAVES Diapositivaslucerito100% (1)
- Objetos A ObservarDocumento17 páginasObjetos A ObservarAmanda Del LagoAinda não há avaliações
- El Carácter Según Los AstrosDocumento12 páginasEl Carácter Según Los Astrosnuevaacropolis_es50% (2)
- Morfologia de La Palma Africana-1Documento5 páginasMorfologia de La Palma Africana-1Ximenita Bel100% (2)
- RassialDocumento2 páginasRassialRandall FlaggAinda não há avaliações
- Septima Clase - EstratigrafíaDocumento84 páginasSeptima Clase - EstratigrafíadannyAinda não há avaliações