Você também pode gostar
- Womens Health Dental School Curriculum 2012Documento81 páginasWomens Health Dental School Curriculum 2012Sex & Gender Women's Health CollaborativeAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- ICSIDocumento1 páginaICSISex & Gender Women's Health CollaborativeAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Integrating Gender Into Health Curriculum For Health ProfessionasDocumento39 páginasIntegrating Gender Into Health Curriculum For Health ProfessionasSex & Gender Women's Health CollaborativeAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- HTN and WomenDocumento10 páginasHTN and WomenSex & Gender Women's Health CollaborativeAinda não há avaliações
- Gender Whats The DifferenceDocumento5 páginasGender Whats The DifferenceSex & Gender Women's Health CollaborativeAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Framework For SexAndGenderDocumento1 páginaFramework For SexAndGenderSex & Gender Women's Health CollaborativeAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- EvidenceDocumento2 páginasEvidenceSex & Gender Women's Health CollaborativeAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- ConsensusStatement For Pregnancy and EpilepsyDocumento1 páginaConsensusStatement For Pregnancy and EpilepsySex & Gender Women's Health CollaborativeAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- CVD UpdateDocumento1 páginaCVD UpdateSex & Gender Women's Health CollaborativeAinda não há avaliações
- Journal of The American College of Cardiology, Volume 43, Issue 5, Pages 900-921Documento1 páginaJournal of The American College of Cardiology, Volume 43, Issue 5, Pages 900-921Sex & Gender Women's Health CollaborativeAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- LEASE CONTRACT Taytay Residentialhouse Kei Inagaki Nena TrusaDocumento6 páginasLEASE CONTRACT Taytay Residentialhouse Kei Inagaki Nena TrusaJaime GonzalesAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Learning Competencies: Table of SpecificationDocumento2 páginasLearning Competencies: Table of Specificationyolanda renos0% (1)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Bob Dylan Diskografija Prevodi PesamaDocumento175 páginasBob Dylan Diskografija Prevodi PesamaJolanda NešovićAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Admission Sos 2013-14090513 PDFDocumento21 páginasAdmission Sos 2013-14090513 PDFmanoj31285manojAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Satellite Motion NotesDocumento23 páginasSatellite Motion NotesVarshLok100% (1)
- VIETTELDocumento20 páginasVIETTELSolgrynAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Commentary On The Book of NahumDocumento9 páginasCommentary On The Book of NahumRev Dr Jeffry Camm JP, MIEPR, MISOPAinda não há avaliações
- Comparative Genomics 2 - PART 1Documento31 páginasComparative Genomics 2 - PART 1NnleinomAinda não há avaliações
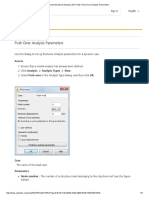
- Robot Structural Analysis 2017 Help - Push Over Analysis ParametersDocumento3 páginasRobot Structural Analysis 2017 Help - Push Over Analysis ParametersJustin MusopoleAinda não há avaliações
- Brochure Bucket SolutionsDocumento20 páginasBrochure Bucket Solutionsmuhammad febri faizinAinda não há avaliações
- Natural Gas DistributionDocumento46 páginasNatural Gas DistributionscribdmisraAinda não há avaliações
- Quad Destroyer - 10 Sets of Squats Workout - Muscle & StrengthDocumento6 páginasQuad Destroyer - 10 Sets of Squats Workout - Muscle & StrengthMouhAinda não há avaliações
- Lesson 4.2 - Operations On Modular ArithmeticDocumento12 páginasLesson 4.2 - Operations On Modular ArithmeticMYLS SHRYNN ELEDAAinda não há avaliações
- Small Incision Cataract SurgeryDocumento249 páginasSmall Incision Cataract SurgeryAillen YovitaAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Neighbor'S Plot - : Sheet No Rev No R0Documento1 páginaNeighbor'S Plot - : Sheet No Rev No R0jibeesh cmAinda não há avaliações
- Fluid Mechanics Formulas Nov 2021Documento6 páginasFluid Mechanics Formulas Nov 2021Benjie MorenoAinda não há avaliações
- Convention Concerning The Protection of The WorldDocumento41 páginasConvention Concerning The Protection of The WorldMonica Ardeleanu100% (1)
- Soal Materi 1 KLS X IntroductionDocumento2 páginasSoal Materi 1 KLS X IntroductionFira AnandaAinda não há avaliações
- Part D - Disclosure and TransparencyDocumento21 páginasPart D - Disclosure and TransparencyMuhammadYudithEddwinaAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Deferred Tax QsDocumento4 páginasDeferred Tax QsDaood AbdullahAinda não há avaliações
- The Brain in Your Pocket Evidence - That - SDocumento8 páginasThe Brain in Your Pocket Evidence - That - Sanwaarft123Ainda não há avaliações
- STAS 111 - Information AgeDocumento20 páginasSTAS 111 - Information AgeMayeee GayosoAinda não há avaliações
- dnd3 Character Sheet STD 105cDocumento2 páginasdnd3 Character Sheet STD 105cGerry MaloneyAinda não há avaliações
- Glide Reflection Folding InstructionsDocumento1 páginaGlide Reflection Folding Instructionsapi-355107616Ainda não há avaliações
- Resume For Singapore Spass Civil EngineerDocumento8 páginasResume For Singapore Spass Civil EngineerArul SD100% (1)
- Price and Output Determination Under OligopolyDocumento26 páginasPrice and Output Determination Under OligopolySangitha Nadar100% (1)
- Wavoo Wajeeha Women's College - Annual Report - 2013-14Documento29 páginasWavoo Wajeeha Women's College - Annual Report - 2013-14kayalonthewebAinda não há avaliações
- Zimbabwe National Code Ccorporate GovernanceDocumento96 páginasZimbabwe National Code Ccorporate GovernanceHerbert NgwaraiAinda não há avaliações
- Clause: Extra Element + Independent Clause Dependent ClauseDocumento1 páginaClause: Extra Element + Independent Clause Dependent ClauseTieng HuangAinda não há avaliações
- 1982 B47 Lehigh County Gunsmithing FamiliesDocumento12 páginas1982 B47 Lehigh County Gunsmithing FamiliesAmr DeabesAinda não há avaliações