Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
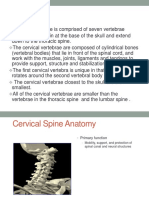
- Emergency Care of Cervical Spine Injuries'Documento38 páginasEmergency Care of Cervical Spine Injuries'Khirman BatoolAinda não há avaliações
- (J. Van Royen Barend, Ben A. C. Dijkmans) Ankylosi PDFDocumento380 páginas(J. Van Royen Barend, Ben A. C. Dijkmans) Ankylosi PDFpremAinda não há avaliações
- Essential Lumbar Special TestsDocumento3 páginasEssential Lumbar Special TestsJesiree DizonAinda não há avaliações
- History of Spine BiomechanicsDocumento13 páginasHistory of Spine BiomechanicsLeopoldo AguayoAinda não há avaliações
- Avian Anatomy: Region or Interramal Space. The Tongue and Related Structures Are Nestled in ThisDocumento36 páginasAvian Anatomy: Region or Interramal Space. The Tongue and Related Structures Are Nestled in Thisdine4ya50% (2)
- Dynamic Deep Squat Lower-Body Kinematics and Considerations Regarding Squat Technique, Load Position, and Heel HeightDocumento10 páginasDynamic Deep Squat Lower-Body Kinematics and Considerations Regarding Squat Technique, Load Position, and Heel HeightThiago LealAinda não há avaliações
- Spondylosis PPTDocumento73 páginasSpondylosis PPTmayuri zanwar100% (2)
- Determining The Age of Fish: BluegillDocumento2 páginasDetermining The Age of Fish: BluegillAndrew BenavidesAinda não há avaliações
- Pott DiseaseDocumento8 páginasPott DiseaseMuhammad IrwantoAinda não há avaliações
- Comparing the Skeletal Systems of Frogs and HumansDocumento15 páginasComparing the Skeletal Systems of Frogs and HumansHannah Nicole TomasAinda não há avaliações
- FIU - Cervical SpineDocumento42 páginasFIU - Cervical Spineamoon12Ainda não há avaliações
- Failed Spinal Anaesthesia Mechanisms, Management and PreventionDocumento6 páginasFailed Spinal Anaesthesia Mechanisms, Management and Preventiondrhemantt1279Ainda não há avaliações
- Neuro LogbookDocumento16 páginasNeuro Logbookbrunosantiago100% (1)
- OKU 5 Orthopaedic Knowledge Update SpineDocumento45 páginasOKU 5 Orthopaedic Knowledge Update SpinePubMed77100% (1)
- HeadDocumento360 páginasHeadfeliciostenio100% (1)
- Certified Interventional Pain Sonologist Exam GuideDocumento19 páginasCertified Interventional Pain Sonologist Exam GuideHisham SalamehAinda não há avaliações
- Minimally Invasive Lumbar Spine SurgeryDocumento29 páginasMinimally Invasive Lumbar Spine SurgerySourabh AlawaAinda não há avaliações
- PGI May 2015 PaperDocumento293 páginasPGI May 2015 PaperRohan RathoreAinda não há avaliações
- MusclesDocumento15 páginasMusclesMeliodas DragneelAinda não há avaliações
- Everything You Need To Know About Supersets For Hypertrophy - Breaking MuscleDocumento4 páginasEverything You Need To Know About Supersets For Hypertrophy - Breaking MuscleEnrique Manuel CerettiAinda não há avaliações
- Orthopaedic ClerkshipDocumento39 páginasOrthopaedic ClerkshippandstutorsAinda não há avaliações
- Spinal ImagingDocumento311 páginasSpinal ImagingguillaumarAinda não há avaliações
- Pathophysiology of pneumothorax and hemothoraxDocumento3 páginasPathophysiology of pneumothorax and hemothoraxAllana RayosAinda não há avaliações
- Scoliosis and Kyphosis2Documento23 páginasScoliosis and Kyphosis2metoAinda não há avaliações
- Marcus Didiano V Dressler, Tuzzolino, USA Hockey, Ontario Junior A Hockey League, OHA, OHF, and Hockey CanadaDocumento15 páginasMarcus Didiano V Dressler, Tuzzolino, USA Hockey, Ontario Junior A Hockey League, OHA, OHF, and Hockey CanadaAnonymous dHerywYZ8pAinda não há avaliações
- ST Marylebone's Paddington Street North Burial Ground: Excavations at Paddington Street, London W1, 2012-13Documento14 páginasST Marylebone's Paddington Street North Burial Ground: Excavations at Paddington Street, London W1, 2012-13MOLA (Museum of London Archaeology)50% (2)
- Iyengar Yoga For Herniated DiscDocumento13 páginasIyengar Yoga For Herniated Discaquabuck100% (8)
- 2018, 840 Pages, Colour Plates, Gures, Tables 25cm: ISBN 938654648-5Documento822 páginas2018, 840 Pages, Colour Plates, Gures, Tables 25cm: ISBN 938654648-5Iryna ZapekaAinda não há avaliações
- Handbook of Bird Biology - CH 4Documento4 páginasHandbook of Bird Biology - CH 4butylacetateAinda não há avaliações
- JAAOS - Volume 05 - Issue 02 March 1997Documento60 páginasJAAOS - Volume 05 - Issue 02 March 1997kenthepaAinda não há avaliações