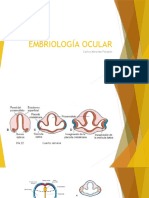
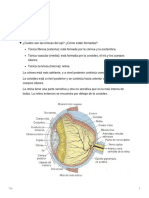
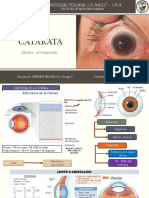
Você também pode gostar
- Toma de Queratometrias y Ley de JavalDocumento7 páginasToma de Queratometrias y Ley de JavalCristopher Contreras100% (1)
- YUEN Las SensacionesDocumento2 páginasYUEN Las SensacionesMarisa Niticia Noya Calviño100% (8)
- Maquillaje ProfesionalDocumento23 páginasMaquillaje Profesionalfantomitas100% (6)
- El Ojo HumanoDocumento18 páginasEl Ojo HumanoJose Manuel Blazquez BlancoAinda não há avaliações
- El Ojo Humano Origen Desarrollo Vision Monocular 6850 CompletoDocumento29 páginasEl Ojo Humano Origen Desarrollo Vision Monocular 6850 CompletoMarion FernandaAinda não há avaliações
- A Retina Es El Órgano Sensorial Del OjoDocumento4 páginasA Retina Es El Órgano Sensorial Del OjoLuisa FERNANDAAinda não há avaliações
- Clase 1 Retina ADocumento6 páginasClase 1 Retina AEma Antonia AlmarzaAinda não há avaliações
- El Ojo Es Un Órgano Sensorial Complejo Que Actúa Como Receptor Del Aparato de La VisiónDocumento5 páginasEl Ojo Es Un Órgano Sensorial Complejo Que Actúa Como Receptor Del Aparato de La VisiónYasna Isabel Molina GuajardoAinda não há avaliações
- RetinaDocumento5 páginasRetinaCarlos EstradaAinda não há avaliações
- Embriogenesis Del Ojo 2014Documento52 páginasEmbriogenesis Del Ojo 2014WalterQunoqYanceOw100% (1)
- Embriologia OcularDocumento49 páginasEmbriologia OcularEsther AlonsoAinda não há avaliações
- 1la RetinaDocumento9 páginas1la RetinaTitto Cedeño VeraAinda não há avaliações
- Anatomia Basica y Fisiologica de La RetinaDocumento11 páginasAnatomia Basica y Fisiologica de La RetinaDigitadores Optica GDAAinda não há avaliações
- Embriología OcularDocumento49 páginasEmbriología OcularLigia Rios100% (1)
- Histologia de Ojo - RetinaDocumento19 páginasHistologia de Ojo - RetinadalianajaelgonzalezAinda não há avaliações
- Fundamentos Físicos de La VisiónDocumento14 páginasFundamentos Físicos de La VisiónLaura RamirezAinda não há avaliações
- Aparato de La VisionDocumento3 páginasAparato de La Visionraul escobarAinda não há avaliações
- Histología Del OjoDocumento5 páginasHistología Del Ojocandela vAinda não há avaliações
- Anatomía Funcional de Retina, Nervio Óptico, Vía Visual y Vías PupilaresDocumento80 páginasAnatomía Funcional de Retina, Nervio Óptico, Vía Visual y Vías Pupilaresjorge perez martinezAinda não há avaliações
- Histo SentidosDocumento7 páginasHisto Sentidoshugo rojasAinda não há avaliações
- Anatomía Del OjoDocumento22 páginasAnatomía Del Ojoyargmilin mendozaAinda não há avaliações
- Tema XXI Desarrollo de Ojos y OidosDocumento9 páginasTema XXI Desarrollo de Ojos y OidosKairich V SojoAinda não há avaliações
- Compendio Oftalmologia RetinaDocumento127 páginasCompendio Oftalmologia Retinamauricioyapur100% (2)
- Embriología Del Globo OcularDocumento8 páginasEmbriología Del Globo OcularYorgenis Nuñez100% (1)
- Embriologia Del Globo OcularDocumento11 páginasEmbriologia Del Globo OcularKaroline Santana100% (2)
- Embriología Del Ojo - VALERIANODocumento6 páginasEmbriología Del Ojo - VALERIANOJosue ValerianoAinda não há avaliações
- 4 Histología de La Visión 1Documento22 páginas4 Histología de La Visión 1cristianoAinda não há avaliações
- RetinaDocumento4 páginasRetinaElainne AlfaroAinda não há avaliações
- 10 Capas de La RetinaDocumento2 páginas10 Capas de La RetinaNaeli ChampoAinda não há avaliações
- 1 Embriología y Anatomía Del Aparato de La VisiónDocumento41 páginas1 Embriología y Anatomía Del Aparato de La VisiónRenato José Casartelli0% (1)
- EstesiologiaDocumento22 páginasEstesiologiaYurani RojasAinda não há avaliações
- Estructura General Del OjoDocumento5 páginasEstructura General Del OjoMariano GamboaAinda não há avaliações
- Informe de Ojo y OidoDocumento15 páginasInforme de Ojo y OidoKarenVivianaAinda não há avaliações
- Anatomía MicroscópicaDocumento8 páginasAnatomía MicroscópicaConejita MorenoAinda não há avaliações
- Descripción Histológica de Las Túnicas Del Globo OcularDocumento10 páginasDescripción Histológica de Las Túnicas Del Globo OcularGipsi MartinezAinda não há avaliações
- INTRODUCCIÓNDocumento20 páginasINTRODUCCIÓNLorenzo QuintanaAinda não há avaliações
- Embriología OcularDocumento20 páginasEmbriología OcularCarlos NarváezAinda não há avaliações
- Anatomia Ocular Leccion 5Documento46 páginasAnatomia Ocular Leccion 5emilyzavala1143009Ainda não há avaliações
- Capas de La RetinaDocumento19 páginasCapas de La Retinaguisela fuentesAinda não há avaliações
- Cuestionario Ojo OidoDocumento10 páginasCuestionario Ojo OidoPaula EloinaAinda não há avaliações
- Sistema de Los SentidosDocumento15 páginasSistema de Los SentidosRaul GonzalezAinda não há avaliações
- Tarea 3 - Via Visual PDFDocumento30 páginasTarea 3 - Via Visual PDFJaimeSergioVasquezZunigaAinda não há avaliações
- OjoDocumento17 páginasOjoGonzalo ManavellaAinda não há avaliações
- Organo de La VisionDocumento33 páginasOrgano de La Visionjunior pinedaAinda não há avaliações
- Histologia de OjoDocumento20 páginasHistologia de OjoKevin Mercado GarciaAinda não há avaliações
- Anatomía Funcional de Retina, Nervio Óptico, Vía Visual y Vías PupilaresDocumento67 páginasAnatomía Funcional de Retina, Nervio Óptico, Vía Visual y Vías PupilaresDr. jorge perez martinez100% (1)
- Desarrollo Del OidoDocumento5 páginasDesarrollo Del Oidolarapaloma698Ainda não há avaliações
- Embriología Del OjoDocumento8 páginasEmbriología Del OjoCionibel Rosario RodriguezAinda não há avaliações
- Resumen de Ojo EmbriologíaDocumento3 páginasResumen de Ojo EmbriologíaJuan CAinda não há avaliações
- Tema 5 D Ojo - OidoDocumento10 páginasTema 5 D Ojo - OidoANA BELEN MIRANDA PERALTAAinda não há avaliações
- Línea Del Tiempo OntogénesisDocumento13 páginasLínea Del Tiempo OntogénesisAna ToraAinda não há avaliações
- Histologia GeneralDocumento6 páginasHistologia GeneralLuis F Concha Manriquez100% (1)
- Embriología Del Globo Ocular.Documento13 páginasEmbriología Del Globo Ocular.Eduardo EstrellaAinda não há avaliações
- Sentido de La Vista en AnimalesDocumento7 páginasSentido de La Vista en AnimalesLi IzzAinda não há avaliações
- Biologia 3actDocumento4 páginasBiologia 3actRosario HernandezAinda não há avaliações
- Histologia - Ojo Uabp9Documento8 páginasHistologia - Ojo Uabp9Seccion Control de Agencias URIAinda não há avaliações
- Embriología Del OjoDocumento36 páginasEmbriología Del Ojooftalmologia hospital posadasAinda não há avaliações
- Embriología Del OjoDocumento36 páginasEmbriología Del Ojooftalmologia hospital posadasAinda não há avaliações
- Embriología y Anatomía Del OjoDocumento17 páginasEmbriología y Anatomía Del OjoCarmen Sofia Mejia PayvaAinda não há avaliações
- Embriologia Del Ojo Humano. Luis Collado 2-17-0344 Grupo 001Documento14 páginasEmbriologia Del Ojo Humano. Luis Collado 2-17-0344 Grupo 001Luis y PoleAinda não há avaliações
- Anatomia Globo OcularDocumento3 páginasAnatomia Globo OcularFabiel RosadoAinda não há avaliações
- Desarrollo Del OjoDocumento2 páginasDesarrollo Del OjoMarisa Niticia Noya CalviñoAinda não há avaliações
- Eft Tapping AbundanciaDocumento1 páginaEft Tapping AbundanciaMarisa Niticia Noya CalviñoAinda não há avaliações
- Suzanne Powell Dieta DisociadaDocumento2 páginasSuzanne Powell Dieta DisociadaMarisa Niticia Noya Calviño0% (1)
- Clase Lupas y PrismasDocumento23 páginasClase Lupas y Prismasbossmiguel667Ainda não há avaliações
- Anatomía y Funciones Generales Del Órgano SensorialDocumento8 páginasAnatomía y Funciones Generales Del Órgano Sensorialtatiana lozanoAinda não há avaliações
- Funciones ReflejasDocumento19 páginasFunciones ReflejasMiriam Ayde Mamani VargasAinda não há avaliações
- Micro Ce LulaDocumento26 páginasMicro Ce LulaAnatomicAinda não há avaliações
- Propedeutica, Semiologia y Estudios de GabineteDocumento22 páginasPropedeutica, Semiologia y Estudios de GabineteBertha Correu AntunezAinda não há avaliações
- Lentes de Contacto Generalidades y AdaptacionDocumento42 páginasLentes de Contacto Generalidades y Adaptacionpqwoeifjpakou13480% (5)
- IFRS For SMEs StandardDocumento369 páginasIFRS For SMEs StandardElvins AguilarAinda não há avaliações
- Metáforas Sobre La VisiónDocumento2 páginasMetáforas Sobre La Visiónanon_465141044Ainda não há avaliações
- Catarata - Oftalmología ExDocumento12 páginasCatarata - Oftalmología Ex11-MH-HU-GEORGE CRISTIANI MIRANDA MACAVILCAAinda não há avaliações
- Oftalmo Basic CAODocumento16 páginasOftalmo Basic CAORodrigo PabonAinda não há avaliações
- La CabezaDocumento14 páginasLa CabezaMario Junior Zegarra Vásquez100% (1)
- Embriologia Del Ojo - Futura MédicaDocumento30 páginasEmbriologia Del Ojo - Futura Médicafuturamedica100% (14)
- Métodos de Sanación Según Conferencias Del Maestro Petar Danov (Omraam Mikhael Aivanhov) PDFDocumento33 páginasMétodos de Sanación Según Conferencias Del Maestro Petar Danov (Omraam Mikhael Aivanhov) PDFAna Maria Laveaga100% (8)
- Alva AnticonaDocumento9 páginasAlva AnticonaMaría Estéfani AlvaradoAinda não há avaliações
- Estrella MacularDocumento37 páginasEstrella MacularDalibor Razmilic BonillaAinda não há avaliações
- La Presbicia o Presbiopía - National Eye InstituteDocumento3 páginasLa Presbicia o Presbiopía - National Eye Instituterodrigo gomezAinda não há avaliações
- AstigmatismoDocumento12 páginasAstigmatismoEdwin Ricardo Ramírez GallardoAinda não há avaliações
- Salud VisualDocumento8 páginasSalud VisualNairo FonsecaAinda não há avaliações
- Herramientas Tecnológicas 1 Fedenador 2020Documento26 páginasHerramientas Tecnológicas 1 Fedenador 2020Katherine AviilaAinda não há avaliações
- Partes Del Ojo y Su MaquillajeDocumento5 páginasPartes Del Ojo y Su MaquillajeNatalia CarreteroAinda não há avaliações
- 5 Basico - Sistema MuscularDocumento3 páginas5 Basico - Sistema MuscularIvan Vargas BustamanteAinda não há avaliações
- Par Craneal 1 Nervio OlfatorioDocumento2 páginasPar Craneal 1 Nervio OlfatorioANDREA VALENTINA RAMIREZ GARCIAAinda não há avaliações
- Conceptos Básicos de Anatomía y Fisiología ....Documento30 páginasConceptos Básicos de Anatomía y Fisiología ....MaterialFacultad100% (1)
- Árbol GenealógicoDocumento10 páginasÁrbol GenealógicoCESAR DAVID SALTOS SALDARRIAGAAinda não há avaliações
- Biología UD 5-8Documento15 páginasBiología UD 5-8Tito JaimeAinda não há avaliações
- Modulo 2 Introduccion A La Ensenanza y Aprendizaje de La Lectura y Escritura xmKdOLgbDocumento130 páginasModulo 2 Introduccion A La Ensenanza y Aprendizaje de La Lectura y Escritura xmKdOLgbguzman camilaAinda não há avaliações
- Leica M50-M60-M80 UserManual ESDocumento66 páginasLeica M50-M60-M80 UserManual ESarturoAinda não há avaliações
- Anatomia Del Ojo Dr. PomatantaDocumento26 páginasAnatomia Del Ojo Dr. PomatantaClaritzaAinda não há avaliações