Você também pode gostar
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- SUPERHERO Suspension Training ManualDocumento11 páginasSUPERHERO Suspension Training ManualCaleb Leadingham100% (5)
- Guidelines For Forensic Report Writing: Helping Trainees Understand Common PitfallsDocumento54 páginasGuidelines For Forensic Report Writing: Helping Trainees Understand Common PitfallsNorfolk Journal100% (1)
- Power of AttorneyDocumento10 páginasPower of AttorneyRocketLawyer82% (17)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Erich FrommDocumento2 páginasErich FrommTina NavarroAinda não há avaliações
- 812571-006 RevB (GPC3000 Service)Documento270 páginas812571-006 RevB (GPC3000 Service)BPAinda não há avaliações
- Annex 9: Guide To Good Storage Practices For PharmaceuticalsDocumento12 páginasAnnex 9: Guide To Good Storage Practices For PharmaceuticalsRyan Noel Perez de TagleAinda não há avaliações
- 2 Effective Manufacturing ERP MESDocumento17 páginas2 Effective Manufacturing ERP MESm_trang2005100% (2)
- EDC MS5 In-Line Injection Pump: Issue 2Documento57 páginasEDC MS5 In-Line Injection Pump: Issue 2Musharraf KhanAinda não há avaliações
- KhanIzh - FGI Life - Offer Letter - V1 - Signed - 20220113154558Documento6 páginasKhanIzh - FGI Life - Offer Letter - V1 - Signed - 20220113154558Izharul HaqueAinda não há avaliações
- Studovaný Okruh: Physical Therapist Sample Test Questions (G5+)Documento8 páginasStudovaný Okruh: Physical Therapist Sample Test Questions (G5+)AndreeaAinda não há avaliações
- 21 Gene Breast AssayDocumento3 páginas21 Gene Breast AssayAnonymous PKE8zOXAinda não há avaliações
- R Atc OopDocumento92 páginasR Atc OopAnonymous PKE8zOXAinda não há avaliações
- Lecture4 PDFDocumento28 páginasLecture4 PDFAnonymous PKE8zOXAinda não há avaliações
- Bayesian Methods For Support Vector Machines: Evidence and Predictive Class ProbabilitiesDocumento32 páginasBayesian Methods For Support Vector Machines: Evidence and Predictive Class ProbabilitiesAnonymous PKE8zOXAinda não há avaliações
- Principles of Cancer StagingDocumento28 páginasPrinciples of Cancer StagingAnonymous PKE8zOXAinda não há avaliações
- Ni Hms 511528Documento19 páginasNi Hms 511528Anonymous PKE8zOXAinda não há avaliações
- Identifying Potential Cancer Driver Genes by Genomic Data IntegrationDocumento9 páginasIdentifying Potential Cancer Driver Genes by Genomic Data IntegrationAnonymous PKE8zOXAinda não há avaliações
- 1 - Introduction To BioinformaticsDocumento28 páginas1 - Introduction To BioinformaticsAnonymous PKE8zOXAinda não há avaliações
- Day1 Laros RNASeq Galaxy 2012Documento40 páginasDay1 Laros RNASeq Galaxy 2012Anonymous PKE8zOXAinda não há avaliações
- How To Understand The Cell by Breaking It: Network Analysis of Gene Perturbation ScreensDocumento13 páginasHow To Understand The Cell by Breaking It: Network Analysis of Gene Perturbation ScreensAnonymous PKE8zOXAinda não há avaliações
- Soal 2-3ADocumento5 páginasSoal 2-3Atrinanda ajiAinda não há avaliações
- Athletes Who Made Amazing Comebacks After Career-Threatening InjuriesDocumento11 páginasAthletes Who Made Amazing Comebacks After Career-Threatening InjuriesანაAinda não há avaliações
- Depression List of Pleasant ActivitiesDocumento3 páginasDepression List of Pleasant ActivitiesShivani SinghAinda não há avaliações
- RISK MANAGEMENT - Imo Multilingual Glossary On Risk Management (Secretariat)Documento17 páginasRISK MANAGEMENT - Imo Multilingual Glossary On Risk Management (Secretariat)Martin NiAinda não há avaliações
- Ammonium Chloride: Product InformationDocumento2 páginasAmmonium Chloride: Product InformationusamaAinda não há avaliações
- BARCODESDocumento7 páginasBARCODESChitPerRhosAinda não há avaliações
- Tracheo Esophageal FistulaDocumento6 páginasTracheo Esophageal Fistulablast2111Ainda não há avaliações
- Owner'S Manual: Explosion-Proof Motor Mf07, Mf10, Mf13Documento18 páginasOwner'S Manual: Explosion-Proof Motor Mf07, Mf10, Mf13mediacampaigncc24Ainda não há avaliações
- Biology 2 Chapter 4Documento61 páginasBiology 2 Chapter 4Malas Nak TaipAinda não há avaliações
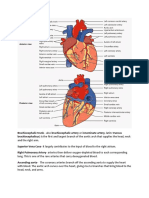
- Brachiocephalic TrunkDocumento3 páginasBrachiocephalic TrunkstephAinda não há avaliações
- 2017 LT4 Wiring DiagramDocumento10 páginas2017 LT4 Wiring DiagramThomasAinda não há avaliações
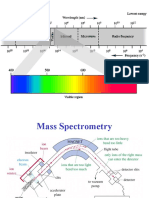
- Tomography: Tomography Is Imaging by Sections or Sectioning Through The Use of AnyDocumento6 páginasTomography: Tomography Is Imaging by Sections or Sectioning Through The Use of AnyJames FranklinAinda não há avaliações
- Mass SpectrometryDocumento49 páginasMass SpectrometryUbaid ShabirAinda não há avaliações
- Capacity Requirement PlanningDocumento17 páginasCapacity Requirement PlanningvamsibuAinda não há avaliações
- Understanding Senior Citizens Outlook of Death Sample FormatDocumento14 páginasUnderstanding Senior Citizens Outlook of Death Sample FormatThea QuibuyenAinda não há avaliações
- 2022.08.09 Rickenbacker ComprehensiveDocumento180 páginas2022.08.09 Rickenbacker ComprehensiveTony WintonAinda não há avaliações
- Phenotype and GenotypeDocumento7 páginasPhenotype and GenotypeIrish Claire Molina TragicoAinda não há avaliações
- Denagard-CTC US Knowledge ReportDocumento4 páginasDenagard-CTC US Knowledge Reportnick224Ainda não há avaliações
- Aircaft Avionics SystemDocumento21 páginasAircaft Avionics SystemPavan KumarAinda não há avaliações
- D05 Directional Control Valves EngineeringDocumento11 páginasD05 Directional Control Valves EngineeringVentas Control HidráulicoAinda não há avaliações