Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- General: Anaemia & Blood Cell Transfusion in Critical IllnessDocumento1 páginaGeneral: Anaemia & Blood Cell Transfusion in Critical IllnessOlfiany Laurenzia PongohAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Thrombomodulin Tumour Biology and PrognosticDocumento4 páginasThrombomodulin Tumour Biology and PrognosticOlfiany Laurenzia PongohAinda não há avaliações
- Neonatal Hyperbilirubinemia and Elevated Liver Enzymes Associated With Thyroid Hormone Deficiency in NeonatesDocumento4 páginasNeonatal Hyperbilirubinemia and Elevated Liver Enzymes Associated With Thyroid Hormone Deficiency in NeonatesOlfiany Laurenzia PongohAinda não há avaliações
- A Case of Aspartate Aminotransferase MacroenzymeDocumento3 páginasA Case of Aspartate Aminotransferase MacroenzymeOlfiany Laurenzia PongohAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Upper Back ExercisesDocumento2 páginasUpper Back ExercisesMarco NgAinda não há avaliações
- Lamaze Seperatingmomandbabyinfographic Final 1Documento1 páginaLamaze Seperatingmomandbabyinfographic Final 1api-264859193Ainda não há avaliações
- Questions From Previous ExamsDocumento6 páginasQuestions From Previous ExamsFaddy OrahaAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- King's Research Portal: 10.1038/nrurol.2016.128Documento28 páginasKing's Research Portal: 10.1038/nrurol.2016.128Rafael MartinsAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Human Walking RoseDocumento248 páginasHuman Walking RosePablo CruzAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Gross Motor Development 6to12yearsDocumento17 páginasGross Motor Development 6to12yearsGul RockzzAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Acute Diarrhea in ChildrenDocumento9 páginasAcute Diarrhea in ChildrenJoao FonsecaAinda não há avaliações
- Jurnal ObturatorDocumento4 páginasJurnal ObturatorEuginia YosephineAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Gamna Gandy BodiesDocumento5 páginasGamna Gandy BodiesTaufik ShidkiAinda não há avaliações
- AffiliationsDocumento9 páginasAffiliationsGurpreet CharaAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Nexium PDFDocumento11 páginasNexium PDFshrikantmsdAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Impetigo in NeonatusDocumento5 páginasImpetigo in Neonatusasep budiyantoAinda não há avaliações
- Independent Nurse Practitioner - Its Future & ScopeDocumento34 páginasIndependent Nurse Practitioner - Its Future & ScopeSanjana Namchoom67% (3)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Dapus InsomniaDocumento3 páginasDapus InsomniaDewi Sartika HarahapAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Fma Ue EnglishDocumento3 páginasFma Ue EnglishAnaa ElleyAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- AP LacrimalDocumento38 páginasAP LacrimalIoana MovileanuAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Genetic Basis of Blood Group Diversity: ReviewDocumento13 páginasGenetic Basis of Blood Group Diversity: ReviewHasna F BidayahAinda não há avaliações
- Naso & Orogastric Tube Placement, Testing & FeedingDocumento10 páginasNaso & Orogastric Tube Placement, Testing & FeedingYwagar Ywagar100% (1)
- Human Gross Anatomy Syllabus-Fall 2010-2011Documento16 páginasHuman Gross Anatomy Syllabus-Fall 2010-2011Zeina Nazih Bou DiabAinda não há avaliações
- Vidyabhivruddhi)Documento71 páginasVidyabhivruddhi)Anonymous ffswriAinda não há avaliações
- Jurnal Dehydration of NewbornDocumento7 páginasJurnal Dehydration of NewbornRSU Aisyiyah PadangAinda não há avaliações
- Contribution by Mark Jackson Precision Ceramics Montclare CaliforniaDocumento3 páginasContribution by Mark Jackson Precision Ceramics Montclare CaliforniaihamihaAinda não há avaliações
- A Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IDocumento28 páginasA Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IJorge Ramírez100% (1)
- Formula FeedsDocumento35 páginasFormula FeedsrandoAinda não há avaliações
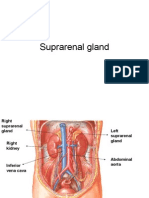
- Suprarenal GlandDocumento19 páginasSuprarenal GlandKay BristolAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- SCNICU Starter PackDocumento7 páginasSCNICU Starter PackALIF HAIQAL MOHD HUSSEINAinda não há avaliações
- Anterior Chamber Angle Assessment Technique - CH 17Documento20 páginasAnterior Chamber Angle Assessment Technique - CH 17riveliAinda não há avaliações
- Closed Fracture HumerusDocumento35 páginasClosed Fracture HumerusEryn Farahin ZainalAinda não há avaliações
- Immediate Care Competency (Baby Care)Documento3 páginasImmediate Care Competency (Baby Care)Louie Jay Ramos100% (1)
- Doctor of Physical Therapy (DPT) : Semester 1Documento5 páginasDoctor of Physical Therapy (DPT) : Semester 1M Irfan Bhutta100% (1)