Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
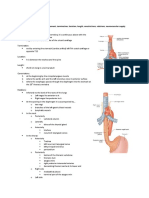
- Anatomy Questions For USMLE (2 of 2)Documento29 páginasAnatomy Questions For USMLE (2 of 2)MedSchoolStuff100% (5)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Virtual EAV: The Electro-Dermal Screening TestDocumento21 páginasVirtual EAV: The Electro-Dermal Screening Testmich100% (1)
- Regulation of Gastric SecretionDocumento18 páginasRegulation of Gastric SecretionSanchit Rao50% (2)
- Visceral Vascular Manipulation, 2011Documento270 páginasVisceral Vascular Manipulation, 2011kkalu_just_funky91% (11)
- DrKapoor BPHDocumento8 páginasDrKapoor BPHRose DeasyAinda não há avaliações
- Jurnal Rabies InggrisDocumento5 páginasJurnal Rabies InggrisArtawa Darma YudaAinda não há avaliações
- Romance de AmorDocumento2 páginasRomance de AmorAlberikus Kwarta BoynoAinda não há avaliações
- IJDFR157Documento11 páginasIJDFR157Alberikus Kwarta BoynoAinda não há avaliações
- Photoshop CS6 Tutorial: Getting Started with Image EditingDocumento27 páginasPhotoshop CS6 Tutorial: Getting Started with Image EditingmulyadiAinda não há avaliações
- KIRBYDocumento5 páginasKIRBYAlberikus Kwarta BoynoAinda não há avaliações
- Romance de AmorDocumento2 páginasRomance de AmorAlberikus Kwarta BoynoAinda não há avaliações
- KIRBYDocumento5 páginasKIRBYAlberikus Kwarta BoynoAinda não há avaliações
- KIRBYDocumento5 páginasKIRBYAlberikus Kwarta BoynoAinda não há avaliações
- KIRBYDocumento5 páginasKIRBYAlberikus Kwarta BoynoAinda não há avaliações
- Regulation of Heart RateDocumento30 páginasRegulation of Heart RateTripat Singh67% (3)
- Embodied Yoga Summit Dr. Arielle Schwartz Vagus Nerve PDFDocumento6 páginasEmbodied Yoga Summit Dr. Arielle Schwartz Vagus Nerve PDFdocerick87100% (1)
- Electromagnetic Fields and It's Effect's On The Hearts Electrical System With Emphasis On The Vagus Nerve A Theoretical Stance, Backed Up by Clinical FactsDocumento3 páginasElectromagnetic Fields and It's Effect's On The Hearts Electrical System With Emphasis On The Vagus Nerve A Theoretical Stance, Backed Up by Clinical FactsegyptarianAinda não há avaliações
- Jurnal NyeriDocumento14 páginasJurnal NyeriIrwan KambuAinda não há avaliações
- Differential Diagnosis BookDocumento101 páginasDifferential Diagnosis BookAmanda PratiwiAinda não há avaliações
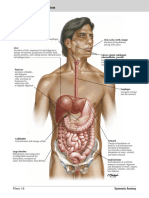
- Esphagus Stomach Duodenum Gross AnatomyDocumento9 páginasEsphagus Stomach Duodenum Gross AnatomyWinchester LoapedAinda não há avaliações
- Stephen W. PorgesDocumento15 páginasStephen W. Porges2rightAinda não há avaliações
- Physioex Lab Report: Pre-Lab Quiz ResultsDocumento3 páginasPhysioex Lab Report: Pre-Lab Quiz Results19-185 Ummu arifah saragihAinda não há avaliações
- Autonomic Nervous System OverviewDocumento16 páginasAutonomic Nervous System OverviewtsyrahmaniAinda não há avaliações
- Martin Torrents DO - Touro College of Osteopathic MedicineDocumento54 páginasMartin Torrents DO - Touro College of Osteopathic MedicineLiv GrahamAinda não há avaliações
- Beginner's GuideDocumento7 páginasBeginner's GuideKitsch nclocksAinda não há avaliações
- Anatomy Quiz 2Documento45 páginasAnatomy Quiz 2Upscaled100% (1)
- A Laboratory Course in Physiology-CannonDocumento337 páginasA Laboratory Course in Physiology-CannonEmilio Jaramillo HolguínAinda não há avaliações
- The Bainbridge and The Reverse Bainbridge.8Documento13 páginasThe Bainbridge and The Reverse Bainbridge.8Archisha KapoorAinda não há avaliações
- Past Papers Fcps Part1Documento1.286 páginasPast Papers Fcps Part1Doc Ahsan50% (4)
- Vagus Nerve Stimulation and Magnet Use - Optimizing Benefits (2009)Documento4 páginasVagus Nerve Stimulation and Magnet Use - Optimizing Benefits (2009)Apróné Török IbolyaAinda não há avaliações
- Skenario 2 Saraf & PerilakuDocumento30 páginasSkenario 2 Saraf & PerilakuPrima Eriawan PutraAinda não há avaliações
- A04 PDFDocumento16 páginasA04 PDFQueen JazAinda não há avaliações
- Dancing With Yin Yang Chap6 ANS LRDocumento34 páginasDancing With Yin Yang Chap6 ANS LRmarianetlaAinda não há avaliações
- 03 Malone Spinal Cord InjuriesDocumento35 páginas03 Malone Spinal Cord InjuriesAnanta MiaAinda não há avaliações
- JV Heart Rate, Heart Sounds PDFDocumento39 páginasJV Heart Rate, Heart Sounds PDFRiya GuptaAinda não há avaliações
- (Netter) Atlas of Human Anatomy. 8° (2023)_3Documento13 páginas(Netter) Atlas of Human Anatomy. 8° (2023)_3ALEXIS SEDERAPAinda não há avaliações
- #1 Points 2 Kool Medic PDFDocumento137 páginas#1 Points 2 Kool Medic PDFSyed Ali Raza NaqviAinda não há avaliações
- Anatomy - Nerves of AbdomenDocumento7 páginasAnatomy - Nerves of AbdomenMuhammad Junaid AnisAinda não há avaliações
- 18 Vessels and Flow DynamicsDocumento57 páginas18 Vessels and Flow Dynamicsraanja2Ainda não há avaliações
- Volume 15, Number 6 June 2011Documento183 páginasVolume 15, Number 6 June 2011Nicolai BabaliciAinda não há avaliações