Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Guidance On The Vigilance System For CE-marked Medical DevicesDocumento4 páginasGuidance On The Vigilance System For CE-marked Medical DevicesbaglamaAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Botanical Society of America, Inc. American Journal of BotanyDocumento6 páginasBotanical Society of America, Inc. American Journal of BotanybaglamaAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- 2 1 6 072016 enDocumento29 páginas2 1 6 072016 enbaglamaAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Ribeiro Et Al 2003Documento7 páginasRibeiro Et Al 2003baglamaAinda não há avaliações
- Studies in The GramineaeDocumento6 páginasStudies in The GramineaebaglamaAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Nea3508 ChernobylDocumento157 páginasNea3508 ChernobylbaglamaAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Deq Ogl Guide Phragmites 204659 7Documento8 páginasDeq Ogl Guide Phragmites 204659 7baglamaAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Int Manual EU28Documento146 páginasInt Manual EU28baglamaAinda não há avaliações
- Ecological Society of AmericaDocumento14 páginasEcological Society of AmericabaglamaAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- w33 1EuroMed 1 Anthemis OretanaDocumento7 páginasw33 1EuroMed 1 Anthemis OretanabaglamaAinda não há avaliações
- Cornicabra TG AG-FoodChem 2004Documento8 páginasCornicabra TG AG-FoodChem 2004baglamaAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Gypsicolous Flora, Conservation and Restoration of Quarries in The Southeast of The Iberian PeninsulaDocumento12 páginasGypsicolous Flora, Conservation and Restoration of Quarries in The Southeast of The Iberian PeninsulabaglamaAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Primer Viaje en Torno Del Globo Written by Antonio Pigafetta. It Was Originally Published in The Year of 1536Documento2 páginasPrimer Viaje en Torno Del Globo Written by Antonio Pigafetta. It Was Originally Published in The Year of 1536Bean BeanAinda não há avaliações
- Julien Freund - A TributeDocumento10 páginasJulien Freund - A TributeJuan Pablo SerraAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- scn615 Classroomgroupactionplan SarahltDocumento3 páginasscn615 Classroomgroupactionplan Sarahltapi-644817377Ainda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Salija, K. (2017) OutlineDocumento9 páginasSalija, K. (2017) OutlineNurina UlfaAinda não há avaliações
- Seventh Day Adventist Quiz by KnowingTheTimeDocumento4 páginasSeventh Day Adventist Quiz by KnowingTheTimeMiiiTheart100% (3)
- 001 Ipack My School BagDocumento38 páginas001 Ipack My School BagBrock JohnsonAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
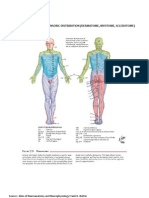
- Dermatome, Myotome, SclerotomeDocumento4 páginasDermatome, Myotome, SclerotomeElka Rifqah100% (3)
- PPH CasestudyDocumento45 páginasPPH CasestudyRona Mae PangilinanAinda não há avaliações
- Ila 0306Documento11 páginasIla 0306Hong ChenAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- 479f3df10a8c0mathsproject QuadrilateralsDocumento18 páginas479f3df10a8c0mathsproject QuadrilateralsAnand PrakashAinda não há avaliações
- Course-Outline EL 102 GenderAndSocietyDocumento4 páginasCourse-Outline EL 102 GenderAndSocietyDaneilo Dela Cruz Jr.Ainda não há avaliações
- The EagleDocumento4 páginasThe EagleJunkoAinda não há avaliações
- Adobe Voice Assessment Tool-FinalDocumento1 páginaAdobe Voice Assessment Tool-Finalapi-268484302Ainda não há avaliações
- The Endless Pursuit of Truth: Subalternity and Marginalization in Post-Neorealist Italian FilmDocumento206 páginasThe Endless Pursuit of Truth: Subalternity and Marginalization in Post-Neorealist Italian FilmPaul MathewAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- (These Are Taken From The Hics Infectious Disease Incident Response Guide Available atDocumento2 páginas(These Are Taken From The Hics Infectious Disease Incident Response Guide Available atDavid MitchellAinda não há avaliações
- Aleister Crowley Astrological Chart - A Service For Members of Our GroupDocumento22 páginasAleister Crowley Astrological Chart - A Service For Members of Our GroupMysticalgod Uidet100% (3)
- Visual Impairments in Young Children: Fundamentals of and Strategies For Enhancing DevelopmentDocumento13 páginasVisual Impairments in Young Children: Fundamentals of and Strategies For Enhancing Developmentfadil ahmadiAinda não há avaliações
- Physics Waves MCQDocumento6 páginasPhysics Waves MCQAyan GhoshAinda não há avaliações
- Art of Data ScienceDocumento159 páginasArt of Data Sciencepratikshr192% (12)
- Invoice Ce 2019 12 IVDocumento8 páginasInvoice Ce 2019 12 IVMoussa NdourAinda não há avaliações
- Academic Calendar 2014 15Documento6 páginasAcademic Calendar 2014 15D Geetha DuraiAinda não há avaliações
- Unit 1 - Identifying A Problem PDFDocumento16 páginasUnit 1 - Identifying A Problem PDFZanko FitnessAinda não há avaliações
- Nutrition and Diet Therapy ExaminationDocumento8 páginasNutrition and Diet Therapy ExaminationIrwan M. Iskober100% (3)
- δ (n) = u (n) - u (n-3) = 1 ,n=0Documento37 páginasδ (n) = u (n) - u (n-3) = 1 ,n=0roberttheivadasAinda não há avaliações
- Anthropology Chapter 2 ADocumento17 páginasAnthropology Chapter 2 AHafiz SaadAinda não há avaliações
- Eco - Module 1 - Unit 3Documento8 páginasEco - Module 1 - Unit 3Kartik PuranikAinda não há avaliações
- Cell Bio1Documento7 páginasCell Bio1AnyaAinda não há avaliações
- Case Study Alliston AssignmentDocumento6 páginasCase Study Alliston AssignmentParamvir SinghAinda não há avaliações
- Servicenow Rest Cheat SheetDocumento3 páginasServicenow Rest Cheat SheetHugh SmithAinda não há avaliações
- Sindhudurg Kokan All Tourism Spot Information WWW - Marathimann.inDocumento54 páginasSindhudurg Kokan All Tourism Spot Information WWW - Marathimann.inMarathi Mann92% (12)