Você também pode gostar
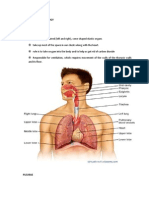
- Vi. Anatomy and Physiology The Lungs StructureDocumento5 páginasVi. Anatomy and Physiology The Lungs StructureJoeven HilarioAinda não há avaliações
- Lab Values: Interpreting Chemistry and Hematology For Adult PatientsDocumento36 páginasLab Values: Interpreting Chemistry and Hematology For Adult PatientsBrian Johnson100% (1)
- Pathophysiology of DiabetesDocumento88 páginasPathophysiology of DiabetesCahya SetiyaAinda não há avaliações
- Insulin Therapy in Type 2 Diabetes MellitusDocumento40 páginasInsulin Therapy in Type 2 Diabetes MellitusEDWIN WIJAYAAinda não há avaliações
- AnaesthesiaDocumento121 páginasAnaesthesiaSandeep Bhangale100% (1)
- MSG Food Addiction in Mice20%10%0%-10%-20%-30%-40%-50%Group IGroup IIGroup IIIGroup IVFIGURE 3. Percentual difference in average liquid consumption from bottles A and B vs. control group IDocumento5 páginasMSG Food Addiction in Mice20%10%0%-10%-20%-30%-40%-50%Group IGroup IIGroup IIIGroup IVFIGURE 3. Percentual difference in average liquid consumption from bottles A and B vs. control group IadriraluAinda não há avaliações
- Medical EmergenciesDocumento115 páginasMedical EmergenciesRamya ReddyAinda não há avaliações
- The Child With Endocrine DysfunctionDocumento5 páginasThe Child With Endocrine Dysfunctionhenny1620Ainda não há avaliações
- Anesthesia NutshellDocumento388 páginasAnesthesia NutshellMuhammad UmerAinda não há avaliações
- Food Poisoning Final Print!Documento3 páginasFood Poisoning Final Print!Joeven HilarioAinda não há avaliações
- Pharmacognostic and Phytochemical Properties of Aloe Vera LinnDocumento5 páginasPharmacognostic and Phytochemical Properties of Aloe Vera LinnazaliaswAinda não há avaliações
- Family Nursing Care PlanDocumento2 páginasFamily Nursing Care PlanJoeven Hilario83% (6)
- Osteosarcoma Case StudyDocumento17 páginasOsteosarcoma Case StudyJoeven HilarioAinda não há avaliações
- Osteosarcoma Case StudyDocumento17 páginasOsteosarcoma Case StudyJoeven HilarioAinda não há avaliações
- Antidiabetic Properties and Mechanism of Action of Gynuria ProcumberDocumento16 páginasAntidiabetic Properties and Mechanism of Action of Gynuria Procumberandirio7486Ainda não há avaliações
- The Effects of Leucaena Leucocephala (LMK) de Wit Seeds On Blood Sugar Levels: An Experimental StudyDocumento4 páginasThe Effects of Leucaena Leucocephala (LMK) de Wit Seeds On Blood Sugar Levels: An Experimental StudyEvi InayatiAinda não há avaliações
- Potent Hypoglycaemic Activity of The Aqueous Extract of Chamaemelum Nobile in Normal and Streptozotocin-Induced Diabetic RatsDocumento7 páginasPotent Hypoglycaemic Activity of The Aqueous Extract of Chamaemelum Nobile in Normal and Streptozotocin-Induced Diabetic RatsMohammedAjebliAinda não há avaliações
- Bengkoang juice effect on diabetes ratsDocumento5 páginasBengkoang juice effect on diabetes ratsGilang Yuka SeptiawanAinda não há avaliações
- Administración 001Documento11 páginasAdministración 001José Luis Haro VeraAinda não há avaliações
- Hypoglicemic EffectDocumento7 páginasHypoglicemic EffectErizal Dwi HandokoAinda não há avaliações
- Effectivity of Lumbrokinase in Reducing Proteinuria On Patients With Diabetic Kidney DiseaseDocumento11 páginasEffectivity of Lumbrokinase in Reducing Proteinuria On Patients With Diabetic Kidney Diseasevedora angeliaAinda não há avaliações
- DiscussionDocumento3 páginasDiscussionMaria Luisa CornelioAinda não há avaliações
- Antidiabetic Effect of Syzygium Cumini L. Seed On Type 2 Diabetic RatsDocumento8 páginasAntidiabetic Effect of Syzygium Cumini L. Seed On Type 2 Diabetic RatsLavanya Priya Sathyan100% (1)
- Momordica Charantia Scientific Paper 16Documento1 páginaMomordica Charantia Scientific Paper 16arjunismeAinda não há avaliações
- 9873 27543 1 PBDocumento5 páginas9873 27543 1 PBManjubashini Leo SelvarajahAinda não há avaliações
- Pengaruh Pemberian Ekstrak Buah Labu Siam (Sechium Edule) Terhadap Penurunan Kadar Glukosa Darah Tikus Wistar Yang Diinduksi AloksanDocumento16 páginasPengaruh Pemberian Ekstrak Buah Labu Siam (Sechium Edule) Terhadap Penurunan Kadar Glukosa Darah Tikus Wistar Yang Diinduksi AloksanAlfi FarmAinda não há avaliações
- Anti-Hyperglycemic and Anti-Hyperlipidemic Effects of Guava Leaf ExtractDocumento17 páginasAnti-Hyperglycemic and Anti-Hyperlipidemic Effects of Guava Leaf ExtractMahendrakumar ManiAinda não há avaliações
- Ajptsp 2007 123 129Documento7 páginasAjptsp 2007 123 129Sri HazizahAinda não há avaliações
- Loss of Early Insulin Secretion Leads To Postprandial HyperglycaemiaDocumento7 páginasLoss of Early Insulin Secretion Leads To Postprandial HyperglycaemiawahyuniAinda não há avaliações
- Glucophage (Metformin Hydrochloride) Tablets Glucophage XR (Metformin Hydrochloride) Extended-Release TabletsDocumento35 páginasGlucophage (Metformin Hydrochloride) Tablets Glucophage XR (Metformin Hydrochloride) Extended-Release TabletsSity DaliAinda não há avaliações
- Vinegar Consumption Increases Insulin-Stimulated GlucoseDocumento7 páginasVinegar Consumption Increases Insulin-Stimulated GlucoseArielLunaAinda não há avaliações
- Effect of pumpkin-based enteral formula on postprandial blood glucose in diabetic ratsDocumento7 páginasEffect of pumpkin-based enteral formula on postprandial blood glucose in diabetic ratsazzahwaauroraAinda não há avaliações
- Association Between Polymorphisms of OCT1 and Metabolic Response To Metformin in Women With Polycystic Ovary Syndrome.Documento10 páginasAssociation Between Polymorphisms of OCT1 and Metabolic Response To Metformin in Women With Polycystic Ovary Syndrome.Fahmi SidiqAinda não há avaliações
- Hypoglycemic and Antioxidant Activities of Balinese Purple Sweet Potato (Ipomoea Batatas L) in Induced-Diabetic RatsDocumento6 páginasHypoglycemic and Antioxidant Activities of Balinese Purple Sweet Potato (Ipomoea Batatas L) in Induced-Diabetic RatsAlfira Naya ZakiyahAinda não há avaliações
- EN Antidiabetic Activity of Sambiloto ExtraDocumento4 páginasEN Antidiabetic Activity of Sambiloto ExtraHasna AthiyahAinda não há avaliações
- Jcmah MS Id 555754Documento6 páginasJcmah MS Id 555754MOHAMMAD OVAISAinda não há avaliações
- Effect of Tomato and Red Guava Juice On Blood Glucose Level in Overweight WomanDocumento6 páginasEffect of Tomato and Red Guava Juice On Blood Glucose Level in Overweight Womanmito kondriaAinda não há avaliações
- Jps R 07021505Documento3 páginasJps R 07021505Oji IfeanyiAinda não há avaliações
- Bhat 2011Documento9 páginasBhat 2011nadin nAinda não há avaliações
- 2022 Anti Dia Be TecDocumento15 páginas2022 Anti Dia Be TecHilda SarahAinda não há avaliações
- PineappleDocumento6 páginasPineappleNatasha Sosa PaidaAinda não há avaliações
- Metformin TimingDocumento11 páginasMetformin Timinglakshminivas PingaliAinda não há avaliações
- Palatinose - Second Meal Effect PDFDocumento7 páginasPalatinose - Second Meal Effect PDFDana ChavezAinda não há avaliações
- APJTBMPublicationDocumento7 páginasAPJTBMPublicationsiti zuhri ramdaniAinda não há avaliações
- Cytotec® Misoprostol TabletsDocumento12 páginasCytotec® Misoprostol Tabletssuhandi71Ainda não há avaliações
- Nutrients: Use of An Extract of Annona Muricata Linn To Prevent High-Fat Diet Induced Metabolic Disorders in C57BLDocumento22 páginasNutrients: Use of An Extract of Annona Muricata Linn To Prevent High-Fat Diet Induced Metabolic Disorders in C57BLjenegneAinda não há avaliações
- MisoprostolDocumento11 páginasMisoprostolMaulidya FakhrulAinda não há avaliações
- Smith 2011Documento7 páginasSmith 2011sari putriAinda não há avaliações
- Jurnal EkstraksiDocumento5 páginasJurnal Ekstraksiandini fasaAinda não há avaliações
- 1 SMDocumento7 páginas1 SMChemayanti SurbaktiAinda não há avaliações
- 2016 - R, Ardiaria PDFDocumento9 páginas2016 - R, Ardiaria PDFJovita AureliaAinda não há avaliações
- Onion Peel Extracts Ameliorate Hyperglycemia and Insulin Resistance in High Fat Diet/ Streptozotocin-Induced Diabetic RatsDocumento8 páginasOnion Peel Extracts Ameliorate Hyperglycemia and Insulin Resistance in High Fat Diet/ Streptozotocin-Induced Diabetic RatsRizky Agustian HadiAinda não há avaliações
- Ginger PublishedDocumento8 páginasGinger PublishedFatimoh AbdulsalamAinda não há avaliações
- en Application of Magnetics Method To MappiDocumento7 páginasen Application of Magnetics Method To MappiNda MiraAinda não há avaliações
- Black Soybean Extract Reduces Blood Glucose in Diabetic RatsDocumento7 páginasBlack Soybean Extract Reduces Blood Glucose in Diabetic RatspryankaAinda não há avaliações
- Mitigating Effects of Vildagliptin in Experimental Diabetes With Metabolic SyndromeDocumento9 páginasMitigating Effects of Vildagliptin in Experimental Diabetes With Metabolic Syndromerajesh sumanAinda não há avaliações
- Gymnema Sylvestre Scientific Paper 22Documento1 páginaGymnema Sylvestre Scientific Paper 22arjunismeAinda não há avaliações
- Antidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusDocumento12 páginasAntidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusAgus FakhrudinAinda não há avaliações
- 1.16 Evaluation of Hypoglycemic Effect of Lagerstroemia Speciosa (Banaba) Leaf Extract in Alloxan Induced Diabetic RabbitsDocumento6 páginas1.16 Evaluation of Hypoglycemic Effect of Lagerstroemia Speciosa (Banaba) Leaf Extract in Alloxan Induced Diabetic RabbitsNikole PetersonAinda não há avaliações
- Evaluation of Medicinal Effects of Gynura Procumbens Leave Extracts On Oxidative Glycemic Lipidomics and Enzymatic ProfiDocumento7 páginasEvaluation of Medicinal Effects of Gynura Procumbens Leave Extracts On Oxidative Glycemic Lipidomics and Enzymatic Profimitu afrinAinda não há avaliações
- 1933 3771 1 SM PDFDocumento16 páginas1933 3771 1 SM PDFFaridaAinda não há avaliações
- Emerging Treatments Improve Diabetes ControlDocumento10 páginasEmerging Treatments Improve Diabetes ControlAyman AleemAinda não há avaliações
- Jpfarmasigg 160035Documento6 páginasJpfarmasigg 160035nikomang ayuAinda não há avaliações
- Beras Coklat (Imam Et Al)Documento18 páginasBeras Coklat (Imam Et Al)monica lestaryAinda não há avaliações
- Imam Et Al 2012Documento18 páginasImam Et Al 2012monica lestaryAinda não há avaliações
- Metformin Ameliorates Diabetes With Metabolic Syndrome Induced Changes in Experimental RatsDocumento11 páginasMetformin Ameliorates Diabetes With Metabolic Syndrome Induced Changes in Experimental RatsGuadalupe Hernandez EquihuaAinda não há avaliações
- 2015 Article 45Documento9 páginas2015 Article 45Dr. Kaushal Kishor SharmaAinda não há avaliações
- Diabetes MelitusDocumento30 páginasDiabetes MelitusSaskiaAinda não há avaliações
- Parisa 201ertyutuer. 1246 012034Documento6 páginasParisa 201ertyutuer. 1246 012034何小霞Ainda não há avaliações
- Uji Aktivitas Antidiabetes Ekstrak Etanol Buah Mengkudu (Morinda Citrifolia L.)Documento8 páginasUji Aktivitas Antidiabetes Ekstrak Etanol Buah Mengkudu (Morinda Citrifolia L.)Use LessAinda não há avaliações
- Terminalia Tomentosa Research DiabetesDocumento4 páginasTerminalia Tomentosa Research DiabetesErshad Shafi AhmedAinda não há avaliações
- Research Article: Antidiabetic Activity of Ethanolic Extract of Hyperglycaemic RatsDocumento6 páginasResearch Article: Antidiabetic Activity of Ethanolic Extract of Hyperglycaemic RatsUmmu Ghodah ZurrahmiAinda não há avaliações
- Manage pain and inflammation with corticosteroid prednisoneDocumento5 páginasManage pain and inflammation with corticosteroid prednisoneJoeven HilarioAinda não há avaliações
- Motives of the Spanish Conquistadors: God, Glory, and GoldDocumento11 páginasMotives of the Spanish Conquistadors: God, Glory, and GoldJoeven HilarioAinda não há avaliações
- Health Teachings: Post Turbt OperationDocumento2 páginasHealth Teachings: Post Turbt OperationJoeven HilarioAinda não há avaliações
- Post-Surgical Pain Assessment and ManagementDocumento2 páginasPost-Surgical Pain Assessment and ManagementJoeven HilarioAinda não há avaliações
- Historical Background and Trends in The Treatment of Infectious DiseasesDocumento50 páginasHistorical Background and Trends in The Treatment of Infectious DiseasesJoeven HilarioAinda não há avaliações
- Mitral Stenosis.. LatestDocumento41 páginasMitral Stenosis.. LatestJoeven HilarioAinda não há avaliações
- 02 TheraphyDocumento2 páginas02 TheraphyJoeven HilarioAinda não há avaliações
- Motives of the Spanish Conquistadors: God, Glory, and GoldDocumento11 páginasMotives of the Spanish Conquistadors: God, Glory, and GoldJoeven HilarioAinda não há avaliações
- Nursing MGT Oral Cav CancerDocumento14 páginasNursing MGT Oral Cav CancerJoeven HilarioAinda não há avaliações
- Vi. Anatomy and PhysiologyDocumento13 páginasVi. Anatomy and PhysiologyJoeven HilarioAinda não há avaliações
- Ke MarcDocumento3 páginasKe MarcJoeven HilarioAinda não há avaliações
- Nursing Care Plan for Hypertension ManagementDocumento3 páginasNursing Care Plan for Hypertension ManagementJoeven HilarioAinda não há avaliações
- BibliographyDocumento3 páginasBibliographyJoeven HilarioAinda não há avaliações
- CrisisDocumento3 páginasCrisisJoeven HilarioAinda não há avaliações
- NCP - FeverDocumento2 páginasNCP - FeverJoeven HilarioAinda não há avaliações
- Liver Cancer IntroDocumento26 páginasLiver Cancer IntroJoeven HilarioAinda não há avaliações
- German MeaslesDocumento16 páginasGerman MeaslesJoeven HilarioAinda não há avaliações
- Drugstudy Last DutyDocumento5 páginasDrugstudy Last DutyJoeven HilarioAinda não há avaliações
- Drugstudy Last DutyDocumento5 páginasDrugstudy Last DutyJoeven HilarioAinda não há avaliações
- Vi. Anatomy and PhysiologyDocumento13 páginasVi. Anatomy and PhysiologyJoeven HilarioAinda não há avaliações
- NCP 2 LykaDocumento1 páginaNCP 2 LykaJoeven HilarioAinda não há avaliações
- Fluorescein Angiography 1Documento2 páginasFluorescein Angiography 1Joeven HilarioAinda não há avaliações
- Lung CancerDocumento20 páginasLung CancerJoeven HilarioAinda não há avaliações
- Health Educ.11Documento5 páginasHealth Educ.11Joeven HilarioAinda não há avaliações
- DM Before and After OperateDocumento5 páginasDM Before and After OperateNararto PrijogoAinda não há avaliações
- Ujian PutriDocumento190 páginasUjian PutriDokterndeso ChannelAinda não há avaliações
- Accu Chek - Guide Me Blood Glucose Meter User Manual v2bDocumento140 páginasAccu Chek - Guide Me Blood Glucose Meter User Manual v2bWao ZapataAinda não há avaliações
- Dr. D.N. Prastowo, SP - PD: Bagian Penyakit Dalam Rs. Hana Charitas - Arga Makmur Bengkulu 2017Documento45 páginasDr. D.N. Prastowo, SP - PD: Bagian Penyakit Dalam Rs. Hana Charitas - Arga Makmur Bengkulu 2017nanikAinda não há avaliações
- ICU Insulin Infusion ProtocolDocumento4 páginasICU Insulin Infusion ProtocolWeeranan Sueb-LaAinda não há avaliações
- New Microsoft Word DocumentDocumento6 páginasNew Microsoft Word DocumentKawther El-FikyAinda não há avaliações
- Bioassay of Insulin Injection: Rabbit and Mouse MethodsDocumento12 páginasBioassay of Insulin Injection: Rabbit and Mouse Methodsm ahsanAinda não há avaliações
- Complications of Parenteral NutritionDocumento24 páginasComplications of Parenteral NutritionAdrián L. VázquezAinda não há avaliações
- Insulin As PartDocumento3 páginasInsulin As PartRezaAinda não há avaliações
- Presentation Indonesia Sep 8 2022Documento88 páginasPresentation Indonesia Sep 8 2022yuni chanAinda não há avaliações
- A Step-By-Step Guide To Using Your Lantus Solostar PenDocumento5 páginasA Step-By-Step Guide To Using Your Lantus Solostar Penraval anil.uAinda não há avaliações
- Litchi, The Unlucky Fruit What Does The Evidence SayDocumento18 páginasLitchi, The Unlucky Fruit What Does The Evidence SaySaujanyaAinda não há avaliações
- Hypoglycemic effects of herbal formulations and gliclazide on diabetesDocumento7 páginasHypoglycemic effects of herbal formulations and gliclazide on diabetesNova RosalinaAinda não há avaliações
- Makalah FarmakoepidemiologiDocumento24 páginasMakalah FarmakoepidemiologiAkbar NugrahaAinda não há avaliações
- Study Guide Blood Glucose MonitoringDocumento4 páginasStudy Guide Blood Glucose MonitoringDan Dan ManaoisAinda não há avaliações
- Med DRADocumento48 páginasMed DRADr-Jagadeesh Mangamoori100% (1)
- Anesthesia For Patients With Diabetes Mellitus - UpToDateDocumento31 páginasAnesthesia For Patients With Diabetes Mellitus - UpToDateEduardoAinda não há avaliações
- Tuvalu Standard Treatment GuidelinesDocumento401 páginasTuvalu Standard Treatment GuidelinesportosinAinda não há avaliações
- HahayysDocumento30 páginasHahayys2BGrp3Plaza, Anna MaeAinda não há avaliações
- Perioperative Management of The Diabetic PatientDocumento9 páginasPerioperative Management of The Diabetic PatientFauzi Abdillah SusmanAinda não há avaliações
- 7396 PDFDocumento7 páginas7396 PDFFranz BackenbauerAinda não há avaliações
- Artificial Intelligence in Medicine: Virginie Felizardo, Nuno M. Garcia, Nuno Pombo, Imen MegdicheDocumento15 páginasArtificial Intelligence in Medicine: Virginie Felizardo, Nuno M. Garcia, Nuno Pombo, Imen MegdicheShujian ZhaoAinda não há avaliações