Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Narcissist ChecklistDocumento3 páginasNarcissist ChecklistSiddharth Chander60% (5)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Screenwriting 101 - Michael ArndtDocumento2 páginasScreenwriting 101 - Michael ArndtSiddharth Chander100% (2)
- Book of Dynamic Assessment in Practice PDFDocumento421 páginasBook of Dynamic Assessment in Practice PDFkamalazizi100% (1)
- Process of CounsellingDocumento15 páginasProcess of CounsellingSamuel Njenga100% (1)
- White Christian America - The EndDocumento108 páginasWhite Christian America - The EndSiddharth ChanderAinda não há avaliações
- Natya Sastra NotesDocumento7 páginasNatya Sastra NotesSiddharth ChanderAinda não há avaliações
- Magic OriginsDocumento2 páginasMagic OriginsSiddharth ChanderAinda não há avaliações
- Posix BasicsDocumento1 páginaPosix BasicsSiddharth ChanderAinda não há avaliações
- Invictus: William Ernest HenleyDocumento1 páginaInvictus: William Ernest HenleySiddharth ChanderAinda não há avaliações
- TruthDocumento1 páginaTruthSiddharth ChanderAinda não há avaliações
- Radio Network Parameters: Wcdma Ran W19Documento12 páginasRadio Network Parameters: Wcdma Ran W19Chu Quang TuanAinda não há avaliações
- Article 1156 Gives The Civil Code Definition of Obligation, in Its Passive Aspect. Our Law MerelyDocumento11 páginasArticle 1156 Gives The Civil Code Definition of Obligation, in Its Passive Aspect. Our Law MerelyFeir, Alexa Mae C.Ainda não há avaliações
- Need You Now Lyrics: Charles Scott, HillaryDocumento3 páginasNeed You Now Lyrics: Charles Scott, HillaryAl UsadAinda não há avaliações
- Challenges For Omnichannel StoreDocumento5 páginasChallenges For Omnichannel StoreAnjali SrivastvaAinda não há avaliações
- Exercise No.2Documento4 páginasExercise No.2Jeane Mae BooAinda não há avaliações
- Paediatrica Indonesiana: Sumadiono, Cahya Dewi Satria, Nurul Mardhiah, Grace Iva SusantiDocumento6 páginasPaediatrica Indonesiana: Sumadiono, Cahya Dewi Satria, Nurul Mardhiah, Grace Iva SusantiharnizaAinda não há avaliações
- #6 Decision Control InstructionDocumento9 páginas#6 Decision Control InstructionTimothy King LincolnAinda não há avaliações
- Q&A JurisdictionDocumento20 páginasQ&A JurisdictionlucasAinda não há avaliações
- Signalling in Telecom Network &SSTPDocumento39 páginasSignalling in Telecom Network &SSTPDilan TuderAinda não há avaliações
- Ergatividad Del Vasco, Teoría Del CasoDocumento58 páginasErgatividad Del Vasco, Teoría Del CasoCristian David Urueña UribeAinda não há avaliações
- Basic Translation TerminologyDocumento7 páginasBasic Translation TerminologyHeidy BarrientosAinda não há avaliações
- Diploma Pendidikan Awal Kanak-Kanak: Diploma in Early Childhood EducationDocumento8 páginasDiploma Pendidikan Awal Kanak-Kanak: Diploma in Early Childhood Educationsiti aisyahAinda não há avaliações
- Sukhtankar Vaishnav Corruption IPF - Full PDFDocumento79 páginasSukhtankar Vaishnav Corruption IPF - Full PDFNikita anandAinda não há avaliações
- NCPDocumento3 páginasNCPchesca_paunganAinda não há avaliações
- SHS11Q4DLP 21st CentFinalDocumento33 páginasSHS11Q4DLP 21st CentFinalNOEMI DE CASTROAinda não há avaliações
- Re CrystallizationDocumento25 páginasRe CrystallizationMarol CerdaAinda não há avaliações
- How To Develop Innovators: Lessons From Nobel Laureates and Great Entrepreneurs. Innovation EducationDocumento19 páginasHow To Develop Innovators: Lessons From Nobel Laureates and Great Entrepreneurs. Innovation Educationmauricio gómezAinda não há avaliações
- Problem ManagementDocumento33 páginasProblem Managementdhirajsatyam98982285Ainda não há avaliações
- Anindya Anticipatory BailDocumento9 páginasAnindya Anticipatory BailYedlaAinda não há avaliações
- Strategic ManagementDocumento14 páginasStrategic ManagementvishakhaAinda não há avaliações
- Peri Operative Nursing ManagementDocumento19 páginasPeri Operative Nursing ManagementSabina KontehAinda não há avaliações
- Sino-Japanese Haikai PDFDocumento240 páginasSino-Japanese Haikai PDFAlina Diana BratosinAinda não há avaliações
- E1979017519 PDFDocumento7 páginasE1979017519 PDFAnant HatkamkarAinda não há avaliações
- Mezbah Uddin Ahmed (173-017-054) Chapter 11Documento12 páginasMezbah Uddin Ahmed (173-017-054) Chapter 11riftAinda não há avaliações
- MAT2355 Final 2002Documento8 páginasMAT2355 Final 2002bojie_97965Ainda não há avaliações
- 619Documento3 páginas619elektron2010Ainda não há avaliações
- A Guide To Relativity BooksDocumento17 páginasA Guide To Relativity Bookscharles luisAinda não há avaliações
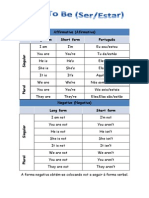
- Affirmative (Afirmativa) Long Form Short Form PortuguêsDocumento3 páginasAffirmative (Afirmativa) Long Form Short Form PortuguêsAnitaYangAinda não há avaliações