Você também pode gostar
- No caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioNo EverandNo caminho certo após uma cirurgia cardíaca: Um guia completo com orientações e recomendações para o pós-operatórioAinda não há avaliações
- 09 - História Da Patologia ResumoDocumento10 páginas09 - História Da Patologia ResumoLarissa Rodrigues VieiraAinda não há avaliações
- O Cérebro Humano: Anatomia, Funções e DoençasDocumento20 páginasO Cérebro Humano: Anatomia, Funções e Doençascamila regis100% (1)
- Câncer de próstata, tratamento, qualidade de vida: como montar este quebra-cabeça?No EverandCâncer de próstata, tratamento, qualidade de vida: como montar este quebra-cabeça?Ainda não há avaliações
- Patologia Clínica e Microbiologia para Técnicos em EnfermagemDocumento26 páginasPatologia Clínica e Microbiologia para Técnicos em EnfermagemEvandrielle FernandesAinda não há avaliações
- Introdução à patologiaDocumento1 páginaIntrodução à patologiaGabriel EliasAinda não há avaliações
- Perguntas de Patologia Que Eu LembroDocumento3 páginasPerguntas de Patologia Que Eu LembroVictoria Pagung0% (1)
- Alterações dos neurotransmissores e implicações clínicas de drogasDocumento57 páginasAlterações dos neurotransmissores e implicações clínicas de drogasClaraAinda não há avaliações
- Introdução à PatologiaDocumento4 páginasIntrodução à PatologiaEdite Do Rosário100% (2)
- Biologia II - CORRIGIDODocumento11 páginasBiologia II - CORRIGIDOKrowl100% (2)
- Dentoalveolar TraumaDocumento7 páginasDentoalveolar TraumaErickDrFb100% (1)
- Fisiopatologia: Bases, Homeostasia e Classificação de DoençasDocumento56 páginasFisiopatologia: Bases, Homeostasia e Classificação de DoençasRaquel FonsecaAinda não há avaliações
- Introdução à PatologiaDocumento26 páginasIntrodução à PatologiaMarcoAinda não há avaliações
- Neuroanatomia Clínica IntrodutóriaDocumento3 páginasNeuroanatomia Clínica IntrodutóriaemanuelAinda não há avaliações
- Sistema Nervoso Autônomo (SNA)Documento38 páginasSistema Nervoso Autônomo (SNA)Fabbio Baldoino100% (1)
- GERIATRIA - Resumo - FisiologiaDocumento35 páginasGERIATRIA - Resumo - FisiologiaLaura A. M. Montouro100% (1)
- Regulação Da Secreção Da ACTHDocumento3 páginasRegulação Da Secreção Da ACTHDanilo BernardoAinda não há avaliações
- DISLIPIDEMIADocumento2 páginasDISLIPIDEMIAbira_tanAinda não há avaliações
- 3aula Fisiologia Sistema Nervoso CentralDocumento39 páginas3aula Fisiologia Sistema Nervoso CentralDeborasSouzaAinda não há avaliações
- Trabalho Farmacologia ReceptoresDocumento10 páginasTrabalho Farmacologia ReceptoresGiulia ArmaneliAinda não há avaliações
- DERMATITE ATÓPICA: REVISÃO SOBRE CAUSAS, SINTOMAS E TRATAMENTODocumento6 páginasDERMATITE ATÓPICA: REVISÃO SOBRE CAUSAS, SINTOMAS E TRATAMENTOKaline AlvesAinda não há avaliações
- Aterosclerose e os vasos sanguíneosDocumento12 páginasAterosclerose e os vasos sanguíneosAna Julia MotaAinda não há avaliações
- Glicocorticóides: propriedades, mecanismos de ação e indicaçõesDocumento75 páginasGlicocorticóides: propriedades, mecanismos de ação e indicaçõesRicardo Alanís100% (1)
- Captura de Tela 2022-11-10 À(s) 01.13.49Documento19 páginasCaptura de Tela 2022-11-10 À(s) 01.13.49Rayane LopesAinda não há avaliações
- Western Blot: Detecção de ProteínasDocumento11 páginasWestern Blot: Detecção de ProteínasKaaio NascimentoAinda não há avaliações
- Introdução à Patologia GeralDocumento19 páginasIntrodução à Patologia GeralAlcsBahiaAinda não há avaliações
- Princípios Da Neurotransmissão Química: Dra. Meiriélle Martins Meneghini PsiquiatriaDocumento55 páginasPrincípios Da Neurotransmissão Química: Dra. Meiriélle Martins Meneghini PsiquiatriaDeyvid Dos AnjosAinda não há avaliações
- Citocinas - ImunologiaDocumento4 páginasCitocinas - ImunologiaAluno - Polyana Aparecida Bisinoto100% (1)
- Resumo Profilaxia HIVDocumento5 páginasResumo Profilaxia HIVAlineAinda não há avaliações
- Qual A Relação Entre Saúde e DoençaDocumento23 páginasQual A Relação Entre Saúde e DoençaCésar OliveiraAinda não há avaliações
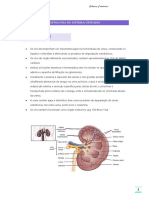
- Anatomia e função dos rins e do sistema urinárioDocumento9 páginasAnatomia e função dos rins e do sistema urinárioSABRINA FURTUNATOAinda não há avaliações
- Febre, icterícia e hepatoesplenomegaliaDocumento2 páginasFebre, icterícia e hepatoesplenomegaliaAmanda Medeiros100% (1)
- Ciclo reprodutivo feminino e gravidezDocumento8 páginasCiclo reprodutivo feminino e gravidezAlynne MoreiraAinda não há avaliações
- Imunidade inata contra fungos e protozoáriosDocumento2 páginasImunidade inata contra fungos e protozoáriosFernanda MotaAinda não há avaliações
- Trabalho DulceDocumento2 páginasTrabalho DulceRamiro ChavesAinda não há avaliações
- Estudo Dirigido Adaptações e Lesoes CelularesDocumento2 páginasEstudo Dirigido Adaptações e Lesoes CelularesJessica BittencourtAinda não há avaliações
- INFLAMAÇÃO E RESPOSTA IMUNEDocumento5 páginasINFLAMAÇÃO E RESPOSTA IMUNEmarisademouraAinda não há avaliações
- Processo de envelhecimento: principais alterações fisiológicasDocumento31 páginasProcesso de envelhecimento: principais alterações fisiológicasEdvan LimaAinda não há avaliações
- Trabalho Sobre NeurotransmissoresDocumento7 páginasTrabalho Sobre NeurotransmissoresRebeca RebouçasAinda não há avaliações
- Epidemiologia conceitosDocumento22 páginasEpidemiologia conceitosFilipe LealAinda não há avaliações
- Cuidados com feridas cirúrgicas e complicaçõesDocumento22 páginasCuidados com feridas cirúrgicas e complicaçõesRenato AvelinoAinda não há avaliações
- RemitDocumento4 páginasRemitIasminAinda não há avaliações
- Síndrome EdwardsDocumento15 páginasSíndrome EdwardsRicardo SantosAinda não há avaliações
- Resposta Neuroendc3b3crina e Metabc3b3lica Ao TraumaDocumento27 páginasResposta Neuroendc3b3crina e Metabc3b3lica Ao TraumaJoão Paulo SacramentoAinda não há avaliações
- A depressão: causas, sintomas e tiposDocumento20 páginasA depressão: causas, sintomas e tiposfmescapeAinda não há avaliações
- Módulo de Proliferação CelularDocumento43 páginasMódulo de Proliferação CelularIsabella GonçalvesAinda não há avaliações
- Western Blot: Técnica de Diagnóstico de Agentes InfecciososDocumento9 páginasWestern Blot: Técnica de Diagnóstico de Agentes InfecciososeguanabaraAinda não há avaliações
- RemitDocumento7 páginasRemitdaysemedAinda não há avaliações
- História Da PatologiaDocumento8 páginasHistória Da PatologiaGiovanna BarbosaAinda não há avaliações
- Obstrução Nasal No Recém-NascidoDocumento7 páginasObstrução Nasal No Recém-NascidoVicente LanaAinda não há avaliações
- Sistema DisgestórioDocumento1 páginaSistema DisgestórioJamille SantosAinda não há avaliações
- Semiotecnica NeurologicaDocumento13 páginasSemiotecnica NeurologicaRainardo PusterAinda não há avaliações
- Alterações hormonais e cardiovasculares no envelhecimentoDocumento21 páginasAlterações hormonais e cardiovasculares no envelhecimentoAriane Gonçalves100% (1)
- Os 5 sentidos: visão, audição, olfato, paladar e tatoDocumento19 páginasOs 5 sentidos: visão, audição, olfato, paladar e tatoLuis Antonio LemesAinda não há avaliações
- Transplantes e Ética MédicaDocumento25 páginasTransplantes e Ética MédicaLuiz AugustoAinda não há avaliações
- Resumo ADAPTAÇÕES - Patologia Geral Aplicada À EnfermagemDocumento15 páginasResumo ADAPTAÇÕES - Patologia Geral Aplicada À EnfermagemAline BitencourtAinda não há avaliações
- Anatomia Patológica SistemáticaDocumento99 páginasAnatomia Patológica SistemáticaMargarida Sequeira MochoAinda não há avaliações
- Conceitos básicos de genéticaDocumento37 páginasConceitos básicos de genéticaLuciana SousaAinda não há avaliações
- Anatomia do Abdome emDocumento67 páginasAnatomia do Abdome emKamila VargasAinda não há avaliações
- Anestesia GeralDocumento62 páginasAnestesia Geralbu_gao2007100% (1)
- Fibrose RetroperitonealDocumento4 páginasFibrose RetroperitonealFisiologia IAinda não há avaliações
- Mesenterico SemiologiaDocumento3 páginasMesenterico SemiologiaFisiologia IAinda não há avaliações
- RoteiroDocumento11 páginasRoteiroFisiologia IAinda não há avaliações
- ABDOME PeritonioDocumento4 páginasABDOME PeritonioFisiologia IAinda não há avaliações
- Anatomia Do Tubo DigestivoDocumento28 páginasAnatomia Do Tubo DigestivoFisiologia IAinda não há avaliações
- Anatomia do Abdome emDocumento67 páginasAnatomia do Abdome emKamila VargasAinda não há avaliações
- Estudo dos ramos sacrais da aorta abdominal do coelhoDocumento10 páginasEstudo dos ramos sacrais da aorta abdominal do coelhoFisiologia IAinda não há avaliações
- Artigo 1Documento6 páginasArtigo 1Fisiologia IAinda não há avaliações
- ABDOME PeritonioDocumento4 páginasABDOME PeritonioFisiologia IAinda não há avaliações
- Informacoes Sobre AnestesiasDocumento5 páginasInformacoes Sobre AnestesiasJulyana BorgesAinda não há avaliações
- Tratamento EndovascularDocumento3 páginasTratamento EndovascularFisiologia IAinda não há avaliações
- Anatomia na prática: testesDocumento103 páginasAnatomia na prática: testesAdriany Carvalho100% (1)
- Breve Historia Da AnestesiaDocumento9 páginasBreve Historia Da AnestesiaFisiologia IAinda não há avaliações
- Anestesia Geral Resumo Teórico 2017 PDFDocumento10 páginasAnestesia Geral Resumo Teórico 2017 PDFFisiologia IAinda não há avaliações
- 05 24 32 Apostila Parasitologia PDFDocumento17 páginas05 24 32 Apostila Parasitologia PDFRafaela de MeloAinda não há avaliações
- Anestesia Geral Resumo Teórico 2017 PDFDocumento10 páginasAnestesia Geral Resumo Teórico 2017 PDFFisiologia IAinda não há avaliações
- Fisiopatologia Da DorDocumento42 páginasFisiopatologia Da DorLeoberto Batista Pereira SobrinhoAinda não há avaliações
- Exerci Cio SDocumento18 páginasExerci Cio SSusana SantosAinda não há avaliações
- Farmacologia dos Anestésicos GeraisDocumento23 páginasFarmacologia dos Anestésicos GeraisFisiologia IAinda não há avaliações
- Tour Cir TraumaDocumento17 páginasTour Cir TraumaFisiologia IAinda não há avaliações
- TSA Topicos EssenciaisDocumento602 páginasTSA Topicos EssenciaisCamila Rodrigues JañaAinda não há avaliações
- Patofisiologia da dor veterináriaDocumento12 páginasPatofisiologia da dor veterináriaFisiologia IAinda não há avaliações
- 3 - Pressão ArterialDocumento4 páginas3 - Pressão ArterialFisiologia IAinda não há avaliações
- O Coração Como BombaDocumento7 páginasO Coração Como BombaFisiologia IAinda não há avaliações
- Eletrofisiologia CardíacaDocumento6 páginasEletrofisiologia CardíacaFisiologia IAinda não há avaliações
- Controle Encefálico Do MovimentoDocumento4 páginasControle Encefálico Do MovimentoFisiologia IAinda não há avaliações
- Apostila PDFDocumento150 páginasApostila PDFGabriel Gaspar Bike Fit100% (2)
- Recursos Estéticos Manuais: Celio Takashi HiguchiDocumento18 páginasRecursos Estéticos Manuais: Celio Takashi HiguchiLorraneAinda não há avaliações
- G1 - CR - Relatório Caso Dor Musculo-EsqueléticaDocumento52 páginasG1 - CR - Relatório Caso Dor Musculo-EsqueléticaInês PinheiroAinda não há avaliações
- Abordagem Fisioterapêutica Na CervicalgiaDocumento74 páginasAbordagem Fisioterapêutica Na CervicalgiaViviane PiresAinda não há avaliações
- Bases Da Terapia Manual OrtopedicaDocumento74 páginasBases Da Terapia Manual OrtopedicaArgleydsson Mendes Durães100% (4)
- Receptores cutâneos e sensibilidade tátilDocumento18 páginasReceptores cutâneos e sensibilidade tátilOdilon Calian0% (1)
- E Book+2021+CompletoDocumento326 páginasE Book+2021+CompletoRafael RochaAinda não há avaliações
- Fisiologia da dor: nocicepção e modulaçãoDocumento35 páginasFisiologia da dor: nocicepção e modulaçãoDEBORA CAPODALIOAinda não há avaliações
- Anamnese, sinais e exame clínicoDocumento17 páginasAnamnese, sinais e exame clínicoCris LavariniAinda não há avaliações
- Seminário de Dor - HM II - A2Documento39 páginasSeminário de Dor - HM II - A2carlao leiAinda não há avaliações
- Apostila Recursos TerapêuticosDocumento57 páginasApostila Recursos TerapêuticosAurilene BelmontAinda não há avaliações
- HarrisonDocumento1.351 páginasHarrisonMaria LimaAinda não há avaliações
- Mobilização Neural: Definição, História e Testes de TensãoDocumento14 páginasMobilização Neural: Definição, História e Testes de TensãoPedro Henrique DoniniAinda não há avaliações
- Trilha Semiologia Da DorDocumento12 páginasTrilha Semiologia Da DorCAMILA EUQUERES PARTATAAinda não há avaliações
- Apitoxina Coleta Composicao Quimica Propriedades BDocumento25 páginasApitoxina Coleta Composicao Quimica Propriedades BMartín NestaAinda não há avaliações
- Resumo - Processos Psicológicos BásicosDocumento21 páginasResumo - Processos Psicológicos BásicosNaylaAinda não há avaliações
- 001 Avaliação SensorialDocumento45 páginas001 Avaliação SensorialAline ChrispimAinda não há avaliações
- Resumo BotoxDocumento5 páginasResumo Botoxnatychristine11Ainda não há avaliações
- Sistema Somatossensorial - Tato, Dor e TemperaturaDocumento56 páginasSistema Somatossensorial - Tato, Dor e TemperaturalaviAinda não há avaliações
- ApostilaDocumento14 páginasApostilaAnderson LimaAinda não há avaliações
- Dor: epidemiologia, fisiopatologia e tratamentoDocumento119 páginasDor: epidemiologia, fisiopatologia e tratamentoMarcelo RomanoAinda não há avaliações
- Caderno - M Dico - Dor. - 171223Documento13 páginasCaderno - M Dico - Dor. - 171223Milson LimaAinda não há avaliações
- Aines 2022.2Documento4 páginasAines 2022.2Marlos Vinícius Batista CoelhoAinda não há avaliações
- Manual DorDocumento120 páginasManual DorRossana RossiAinda não há avaliações
- Apostila Fisiologia Da DORDocumento12 páginasApostila Fisiologia Da DORGabriel Messias100% (1)
- Relatório de Fisiologia sobre Sensação e DorDocumento17 páginasRelatório de Fisiologia sobre Sensação e DorArquimeniaAinda não há avaliações
- Células que sentem dor: nociceptoresDocumento93 páginasCélulas que sentem dor: nociceptoresJoiceRodolphoAinda não há avaliações
- Fisiologia da Dor emDocumento19 páginasFisiologia da Dor emvanessa C.Ainda não há avaliações
- Habilidades sociais de pacientes fibromiálgicasDocumento111 páginasHabilidades sociais de pacientes fibromiálgicasKARIN RIZZIAinda não há avaliações
- Fisiologia Da DorDocumento14 páginasFisiologia Da DorPedro Henrique DoniniAinda não há avaliações