Você também pode gostar
- Especializações Da Membrana PlasmáticaDocumento18 páginasEspecializações Da Membrana PlasmáticaPaula AlmeidaAinda não há avaliações
- Respostas CHGDocumento4 páginasRespostas CHGPablo MonticeliAinda não há avaliações
- Aula 4Documento12 páginasAula 4Aniceto RosarioAinda não há avaliações
- TECIDO EPITELIAL - ResumoDocumento9 páginasTECIDO EPITELIAL - ResumoLetícia S. de CarvalhoAinda não há avaliações
- O conceito e componentes da célulaDocumento8 páginasO conceito e componentes da célulaRoberto Eduardo VieiraAinda não há avaliações
- Citoesqueleto e CitoplasmaDocumento35 páginasCitoesqueleto e CitoplasmaRicardo GomesAinda não há avaliações
- Aula 4 - Biomembranas e OrganelasDocumento56 páginasAula 4 - Biomembranas e OrganelasxisbrjamillyAinda não há avaliações
- Tecido EpitelialDocumento8 páginasTecido EpitelialMaicon Souza LuizAinda não há avaliações
- Citoesqueleto_20231020_164741_0000Documento42 páginasCitoesqueleto_20231020_164741_0000maryemy900Ainda não há avaliações
- Os tecidos epiteliais: características e funçõesDocumento86 páginasOs tecidos epiteliais: características e funçõesBorisAinda não há avaliações
- A estrutura e função da membrana plasmáticaDocumento2 páginasA estrutura e função da membrana plasmáticaMaria Theresa MoreiraAinda não há avaliações
- 1 - Organização Estrutural e Funcional Das CélulasDocumento17 páginas1 - Organização Estrutural e Funcional Das CélulasEduardo MatosAinda não há avaliações
- Guia definitivo sobre CitologiaDocumento65 páginasGuia definitivo sobre CitologiaSuelen ChristineAinda não há avaliações
- Introdução a BioquímicaDocumento6 páginasIntrodução a BioquímicaDeborah RochaAinda não há avaliações
- Fisiopatologia Da Pele: Prof Beatriz Essenfelder BorgesDocumento97 páginasFisiopatologia Da Pele: Prof Beatriz Essenfelder Borgeslii pereiraAinda não há avaliações
- Aula 1Documento16 páginasAula 1Larissa Nacif CanalAinda não há avaliações
- Organelas e estruturas citoplasmáticasDocumento7 páginasOrganelas e estruturas citoplasmáticasEduardo MottaAinda não há avaliações
- Citoesqueleto - SanarflixDocumento12 páginasCitoesqueleto - SanarflixcavalcanathaliaAinda não há avaliações
- Exercícios Citoesqueleto e Núcleo - 2º Período Bio MolDocumento5 páginasExercícios Citoesqueleto e Núcleo - 2º Período Bio MolANJOAinda não há avaliações
- FlipbookDocumento21 páginasFlipbookJohn JohnAinda não há avaliações
- Roteiro de Junção Celular ResolvidoDocumento2 páginasRoteiro de Junção Celular Resolvidothurossi85Ainda não há avaliações
- Matriz extracelular e suas funçõesDocumento13 páginasMatriz extracelular e suas funçõesFabiana RibeiroAinda não há avaliações
- AD2 BiocelDocumento2 páginasAD2 BiocelTaís SantosAinda não há avaliações
- Material CITOLOGIA ORGANELASDocumento10 páginasMaterial CITOLOGIA ORGANELASScaryvanAinda não há avaliações
- Respostas Estudo Dirigido 1 (Com Figurinhas)Documento17 páginasRespostas Estudo Dirigido 1 (Com Figurinhas)RogerMeloAinda não há avaliações
- Células eucariontes: estrutura e funçãoDocumento5 páginasCélulas eucariontes: estrutura e funçãoLucas NevesAinda não há avaliações
- Membrana Plasmatica BioquimDocumento83 páginasMembrana Plasmatica Bioquimflavia douraoAinda não há avaliações
- Resumo de Tecido Epitelial - UFRJDocumento2 páginasResumo de Tecido Epitelial - UFRJhiagoaraujo1999Ainda não há avaliações
- Unid 2Documento23 páginasUnid 2renato bryanAinda não há avaliações
- Os principais componentes do citoesqueletoDocumento8 páginasOs principais componentes do citoesqueletoPaty FernandesAinda não há avaliações
- Membrana CelularDocumento3 páginasMembrana CelularRodrigo KerberAinda não há avaliações
- Resumo Histologia JunqueiraDocumento8 páginasResumo Histologia JunqueiraEmilly EduardaAinda não há avaliações
- 10 - CitoesqueletoDocumento13 páginas10 - CitoesqueletoForwomenAinda não há avaliações
- Citoesqueleto - estruturas e movimentosDocumento7 páginasCitoesqueleto - estruturas e movimentosDanilo TkaczAinda não há avaliações
- Citoesqueleto 2 2019Documento79 páginasCitoesqueleto 2 2019Roberto JunioAinda não há avaliações
- Histologia: 1. Tecido EpitelialDocumento43 páginasHistologia: 1. Tecido EpitelialThaissa MonsoresAinda não há avaliações
- Membrana Plasmática. Digestão IntracelularDocumento27 páginasMembrana Plasmática. Digestão IntracelularBruno PereiraAinda não há avaliações
- Citoesqueleto e movimentos celularesDocumento2 páginasCitoesqueleto e movimentos celularesKeyla SilvaAinda não há avaliações
- Junções celularesDocumento9 páginasJunções celularesRafael AlmeidaAinda não há avaliações
- Organização dos filamentos do citoesqueletoDocumento17 páginasOrganização dos filamentos do citoesqueletoFabiana RibeiroAinda não há avaliações
- Introdução Ao Estudo Da CélulaDocumento25 páginasIntrodução Ao Estudo Da Céluladrifrancas88100% (10)
- Prova Geral 3 - BC180 - RespostasDocumento4 páginasProva Geral 3 - BC180 - RespostasThiago RibasAinda não há avaliações
- Tecidos Epitelial e Conjuntivo PDFDocumento17 páginasTecidos Epitelial e Conjuntivo PDFFlora GuivalaAinda não há avaliações
- Tecido EpitelialDocumento5 páginasTecido EpitelialCamilaAinda não há avaliações
- Wa0030.Documento26 páginasWa0030.Pablo MonticeliAinda não há avaliações
- Turmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143Documento17 páginasTurmadefevereiro-biologia1-Tipos de Células e Membrana Plasmática-12!04!2023-Fb6326a8fa586694572784406618a143bio-luysasantosAinda não há avaliações
- BiocelDocumento9 páginasBiocelmaria joao sargentoAinda não há avaliações
- A Base Celular Da VidaDocumento8 páginasA Base Celular Da VidaRicardo Moura100% (1)
- Célula e Organização da VidaDocumento25 páginasCélula e Organização da VidaAfonso FreireAinda não há avaliações
- Citoesqueleto resumoDocumento6 páginasCitoesqueleto resumoDiana VicenteAinda não há avaliações
- A descoberta de como a mitocôndria regula o cálcio nas célulasDocumento8 páginasA descoberta de como a mitocôndria regula o cálcio nas célulasKarina PingarilhoAinda não há avaliações
- Organelas Celulares e Estruturas Celulares.Documento21 páginasOrganelas Celulares e Estruturas Celulares.Daiane LorenzonAinda não há avaliações
- CitosesqueletoDocumento5 páginasCitosesqueletoGraceAinda não há avaliações
- Aula II - Revisão Sobre Célula Vegetal e As Principais Organelas PDFDocumento34 páginasAula II - Revisão Sobre Célula Vegetal e As Principais Organelas PDFJefferson SauloAinda não há avaliações
- Introdução à estrutura e componentes da célula humanaDocumento15 páginasIntrodução à estrutura e componentes da célula humanaFabio Delfim JuliasseAinda não há avaliações
- Formulario IdososgDocumento1 páginaFormulario IdososgInês LapaAinda não há avaliações
- 4 IndicadoresSaude PDFDocumento33 páginas4 IndicadoresSaude PDFInês LapaAinda não há avaliações
- Weekly Meal Planner PDFDocumento1 páginaWeekly Meal Planner PDFInês LapaAinda não há avaliações
- ESMO ACF Cancro Colorretal Um Guia para o Doente PDFDocumento53 páginasESMO ACF Cancro Colorretal Um Guia para o Doente PDFInês LapaAinda não há avaliações
- 3 MedidasMortalidade PDFDocumento23 páginas3 MedidasMortalidade PDFInês LapaAinda não há avaliações
- ExerciciosMedidasFrequencia PDFDocumento7 páginasExerciciosMedidasFrequencia PDFInês LapaAinda não há avaliações
- 7 EstudosCoorte PDFDocumento23 páginas7 EstudosCoorte PDFInês LapaAinda não há avaliações
- 9 EstudosExperimentais PDFDocumento18 páginas9 EstudosExperimentais PDFInês LapaAinda não há avaliações
- 8 EstudosCaso Controlo PDFDocumento22 páginas8 EstudosCaso Controlo PDFInês LapaAinda não há avaliações
- ExerciciosIndicadoresSaude PDFDocumento3 páginasExerciciosIndicadoresSaude PDFInês LapaAinda não há avaliações
- 5 TiposEstudos PDFDocumento14 páginas5 TiposEstudos PDFInês LapaAinda não há avaliações
- 2 MedidasFrequencia PDFDocumento30 páginas2 MedidasFrequencia PDFInês LapaAinda não há avaliações
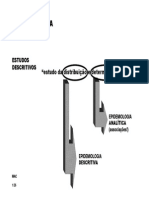
- 6 EstudosDescritivos PDFDocumento26 páginas6 EstudosDescritivos PDFInês LapaAinda não há avaliações
- ExerciciosMedidasMortalidade PDFDocumento11 páginasExerciciosMedidasMortalidade PDFInês LapaAinda não há avaliações
- ExerciciosEstudosAnaliticos PDFDocumento9 páginasExerciciosEstudosAnaliticos PDFInês Lapa0% (1)
- ExerciciosIndicadoresSaude PDFDocumento3 páginasExerciciosIndicadoresSaude PDFInês LapaAinda não há avaliações
- 5FisiologiaSistemaMuscular PDFDocumento92 páginas5FisiologiaSistemaMuscular PDFInês LapaAinda não há avaliações
- 5 TiposEstudos PDFDocumento14 páginas5 TiposEstudos PDFInês LapaAinda não há avaliações
- Sistema Endócrino em 40Documento56 páginasSistema Endócrino em 40Inês Lapa100% (1)
- Miologia PDFDocumento15 páginasMiologia PDFInês LapaAinda não há avaliações
- Notas RD 2011-2012 - MA 2a Frequencia PDFDocumento4 páginasNotas RD 2011-2012 - MA 2a Frequencia PDFInês LapaAinda não há avaliações
- Bioquimica I - Cinetica B W PDFDocumento112 páginasBioquimica I - Cinetica B W PDFInês LapaAinda não há avaliações
- Exercicios Das Praticas MA 2011-2012 PDFDocumento10 páginasExercicios Das Praticas MA 2011-2012 PDFInês LapaAinda não há avaliações
- Sistema Esquelético PDFDocumento20 páginasSistema Esquelético PDFInês LapaAinda não há avaliações
- Sistema Urinário PDFDocumento30 páginasSistema Urinário PDFInês Lapa100% (1)
- Sistema Tegumentar PDFDocumento71 páginasSistema Tegumentar PDFInês LapaAinda não há avaliações
- Anatomia o Sistema ImunitarioDocumento28 páginasAnatomia o Sistema ImunitarioFiá DaminaAinda não há avaliações
- Doença CelíacaDocumento3 páginasDoença CelíacaInês LapaAinda não há avaliações
- Sistema Digestivo Anatomia e FisiologiaDocumento74 páginasSistema Digestivo Anatomia e FisiologiaInês LapaAinda não há avaliações
- Efeito do pH e inibidores na atividade da enzima lacaseDocumento5 páginasEfeito do pH e inibidores na atividade da enzima lacaseInês Lapa100% (1)
- CrescimentoCraniofacialDocumento2 páginasCrescimentoCraniofacialrayanne vinha100% (1)
- Gastroenterologia Essencial. Renato DaniDocumento7 páginasGastroenterologia Essencial. Renato Danimailson eleuterioAinda não há avaliações
- Sopro No CoraçãoDocumento6 páginasSopro No CoraçãoThairone NeryAinda não há avaliações
- Auxiliares Invisíveis ajudam menina e homem em perigoDocumento17 páginasAuxiliares Invisíveis ajudam menina e homem em perigoLucas FernandoAinda não há avaliações
- Dicionário Trilíngue Capovilla - LBS, Libras - YDocumento8 páginasDicionário Trilíngue Capovilla - LBS, Libras - YPriscila Regina Silva100% (4)
- Aula 14 - Microbiologia de CarnesDocumento25 páginasAula 14 - Microbiologia de Carnescarol BrbelvinoAinda não há avaliações
- FluidTherapyGLS en PTDocumento11 páginasFluidTherapyGLS en PTIsabelle PassosAinda não há avaliações
- Doença PeriodontalDocumento24 páginasDoença PeriodontalLydia TavaresAinda não há avaliações
- 9 - Medicina de Mamíferos (Zoo)Documento54 páginas9 - Medicina de Mamíferos (Zoo)Thiago SulzAinda não há avaliações
- LIGA LADO - O Que Pode Causar A Gengivite GranulomatosaDocumento16 páginasLIGA LADO - O Que Pode Causar A Gengivite GranulomatosaTelessaúde AmazonasAinda não há avaliações
- 500 Questoes CTBMF Por Lais AlvesDocumento180 páginas500 Questoes CTBMF Por Lais AlvesTharsis ScaloviskyAinda não há avaliações
- Animais InvertebradosDocumento5 páginasAnimais InvertebradosguilhermeAinda não há avaliações
- Identificação das partes do coraçãoDocumento3 páginasIdentificação das partes do coraçãoLourenço RibeiroAinda não há avaliações
- DESCODIDocumento6 páginasDESCODIAlain JezequelAinda não há avaliações
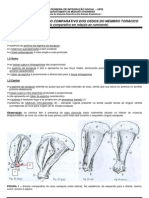
- Comparativo ossos membro torácico animaisDocumento7 páginasComparativo ossos membro torácico animaisMarcélia Almeida100% (2)
- Músculos Do Pescoço - Sistema Muscular - Site Anatomia - Aula Anatomia Humana - Miologia PDFDocumento15 páginasMúsculos Do Pescoço - Sistema Muscular - Site Anatomia - Aula Anatomia Humana - Miologia PDFAnonymous t4FH1JWdoAinda não há avaliações
- Sistema mastigatório: anatomia funcional e componentes anatômicosDocumento16 páginasSistema mastigatório: anatomia funcional e componentes anatômicoskuico2011Ainda não há avaliações
- Manual de apoio ao Curso de Sépsis e Infeção Grave para Médicos (CSIG MedDocumento312 páginasManual de apoio ao Curso de Sépsis e Infeção Grave para Médicos (CSIG MedPedro Rodriguez DiazAinda não há avaliações
- Artrologia AnimalDocumento16 páginasArtrologia AnimalFrancisco Rafael100% (1)
- Como Espantar Afastar Pombos de Casa Telhado PDFDocumento5 páginasComo Espantar Afastar Pombos de Casa Telhado PDFbratoxbrasiliaAinda não há avaliações
- Sistema EndócrinoDocumento37 páginasSistema EndócrinoLaiza BarbinoAinda não há avaliações
- Classificação e estruturas locomotoras de protozoáriosDocumento11 páginasClassificação e estruturas locomotoras de protozoáriosDenner SouzaAinda não há avaliações
- GUIA ABSOLUTO DO DRUIDADocumento4 páginasGUIA ABSOLUTO DO DRUIDASamuel NogueiraAinda não há avaliações
- Resumo SEMIOLOGIA DA PELE E MUCOSAS APARENTESDocumento3 páginasResumo SEMIOLOGIA DA PELE E MUCOSAS APARENTESDário Divino DiasAinda não há avaliações
- Organização da cozinha emDocumento89 páginasOrganização da cozinha emmonica oura100% (2)
- Guia Do Herbívoro Feliz PDFDocumento78 páginasGuia Do Herbívoro Feliz PDFNatália Coltri FernandesAinda não há avaliações
- Albúm Seriado - CópiaDocumento14 páginasAlbúm Seriado - CópiaDanila LorenaAinda não há avaliações
- Aparelho FonadorDocumento9 páginasAparelho FonadorIzadora FrançaAinda não há avaliações
- Exames radiológicos do sistema digestivo e urinárioDocumento73 páginasExames radiológicos do sistema digestivo e urinárioMadeira Luis100% (2)
- Aplicações do Jin Shin JyutsuDocumento4 páginasAplicações do Jin Shin JyutsuMaisa Oliveira100% (3)