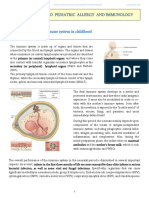
Você também pode gostar
- The Maturing Immune System: Neonatal Immunity: Table 1Documento2 páginasThe Maturing Immune System: Neonatal Immunity: Table 1Vania ParamithaAinda não há avaliações
- Immune System of ChildrenDocumento6 páginasImmune System of Childrenbholasimran93Ainda não há avaliações
- Rol de La Inmunidad Innata en La Infección Neonatal. 2013 PDFDocumento8 páginasRol de La Inmunidad Innata en La Infección Neonatal. 2013 PDFVictor AyalaAinda não há avaliações
- Immunity and Allergy. AsthmaDocumento21 páginasImmunity and Allergy. AsthmaSalman SathooAinda não há avaliações
- INTESTINO FELIZ, ANIMAL SALUDABLE EsDocumento22 páginasINTESTINO FELIZ, ANIMAL SALUDABLE EsurielAinda não há avaliações
- Reproductive Immunology: Basic ConceptsNo EverandReproductive Immunology: Basic ConceptsGil MorAinda não há avaliações
- Vaccination in Early LifeDocumento8 páginasVaccination in Early LifeLeonardo PalenciaAinda não há avaliações
- Neonatal SepsisDocumento22 páginasNeonatal Sepsisjake1014100% (1)
- Immune System - Development and Acquisition of Immunological Competence - 2020Documento8 páginasImmune System - Development and Acquisition of Immunological Competence - 2020Cláudia SilvaAinda não há avaliações
- Immunology in Pregnancy & Coagulation Failure in PregnancyDocumento30 páginasImmunology in Pregnancy & Coagulation Failure in PregnancySrikutty DevuAinda não há avaliações
- Neonatal SepsisDocumento17 páginasNeonatal SepsisDhilla Feroh Kesuma TAinda não há avaliações
- Sepsis Neonatorum UpdatedDocumento87 páginasSepsis Neonatorum Updated'Eduard Laab AlcantaraAinda não há avaliações
- Tutor Guide Astro BoyDocumento10 páginasTutor Guide Astro BoyMaria ArianAinda não há avaliações
- Original Research Paper Paediatric MedicineDocumento4 páginasOriginal Research Paper Paediatric MedicineSujoy DasAinda não há avaliações
- Mol3005 H08Documento13 páginasMol3005 H08DenizDerenAinda não há avaliações
- Infecciones NeonatalesDocumento23 páginasInfecciones NeonatalesAlejandra MedinaAinda não há avaliações
- Antiinfective Properties of Human MilkDocumento6 páginasAntiinfective Properties of Human Milkkiko arrojasAinda não há avaliações
- Chapter 2 - Immunological Changes in Pregnancy and - 2021 - Covid 19 InfectionsDocumento15 páginasChapter 2 - Immunological Changes in Pregnancy and - 2021 - Covid 19 InfectionsWalter MendozaAinda não há avaliações
- Fast Facts: Managing immune-related Adverse Events in Oncology: Early recognition, prompt intervention, effective managementNo EverandFast Facts: Managing immune-related Adverse Events in Oncology: Early recognition, prompt intervention, effective managementAinda não há avaliações
- Immune Responses in NeonatesDocumento26 páginasImmune Responses in NeonatesCony GSAinda não há avaliações
- Desarrollo Temprano Del Sistema Inmune PDFDocumento21 páginasDesarrollo Temprano Del Sistema Inmune PDFVictor AyalaAinda não há avaliações
- 2 Pa Tho Physiology of HIVDocumento8 páginas2 Pa Tho Physiology of HIVLeepin TanAinda não há avaliações
- 2 Pathophysiology of HIVDocumento8 páginas2 Pathophysiology of HIVRaga ManduaruAinda não há avaliações
- Non-inflammatory immunology: An introduction to the immune system and its pathologiesNo EverandNon-inflammatory immunology: An introduction to the immune system and its pathologiesAinda não há avaliações
- EbolaDocumento2 páginasEbolaJosipa ĆevidAinda não há avaliações
- AIDS and ImmunodeficiencyDocumento12 páginasAIDS and ImmunodeficiencyRashed ShatnawiAinda não há avaliações
- Innate Immunity: Fig 1 Table IDocumento9 páginasInnate Immunity: Fig 1 Table IforeveraldyAinda não há avaliações
- Imunology PregnancyDocumento9 páginasImunology PregnancyimeldafitriAinda não há avaliações
- Bookshelf: Chapter 8specific Acquired ImmunityDocumento12 páginasBookshelf: Chapter 8specific Acquired ImmunityBlake KamminAinda não há avaliações
- Peran Imunitas Seluler Pada Ibu Hamil: Arjilio T. Z. Runtukahu, Sylvia R. Marunduh, Hedison PoliiDocumento7 páginasPeran Imunitas Seluler Pada Ibu Hamil: Arjilio T. Z. Runtukahu, Sylvia R. Marunduh, Hedison PoliiPutri MaesarrahAinda não há avaliações
- 04 Intravenous Immune Globulin Clinical Applications in The NewbornDocumento11 páginas04 Intravenous Immune Globulin Clinical Applications in The NewbornMorales Eli Pediatra100% (1)
- Anna Rozaliyani: Department of Parasitology-Faculty of Medicine, Universitas IndonesiaDocumento27 páginasAnna Rozaliyani: Department of Parasitology-Faculty of Medicine, Universitas Indonesiainka.elseAinda não há avaliações
- Fetal-Specific CD8+ Cytotoxic T Cell Responses Develop During Normal Human Pregnancy and Exhibit Broad Functional CapacityDocumento10 páginasFetal-Specific CD8+ Cytotoxic T Cell Responses Develop During Normal Human Pregnancy and Exhibit Broad Functional Capacity石一Ainda não há avaliações
- Nutrients: Diet and Immune FunctionDocumento9 páginasNutrients: Diet and Immune FunctionjenniAinda não há avaliações
- Preclinical Immunology and Microbiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1No EverandPreclinical Immunology and Microbiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1Nota: 5 de 5 estrelas5/5 (1)
- Micronutrients Influencing The Immune Response in Leprosy: RevisiónDocumento11 páginasMicronutrients Influencing The Immune Response in Leprosy: RevisiónSherlocknovAinda não há avaliações
- Allergy: Back To IndexDocumento8 páginasAllergy: Back To IndexjnsenguptaAinda não há avaliações
- The Germinal Center Is An Important Site ForDocumento11 páginasThe Germinal Center Is An Important Site ForGopika SureshAinda não há avaliações
- Gut: Key Element On Immune System Regulation: Review - Human and Animal HealthDocumento14 páginasGut: Key Element On Immune System Regulation: Review - Human and Animal HealthyutefupAinda não há avaliações
- J Immunol 2010 Mueller 2182 90Documento10 páginasJ Immunol 2010 Mueller 2182 90winsarkarAinda não há avaliações
- Peran Imunitas Seluler Pada Ibu HamilDocumento8 páginasPeran Imunitas Seluler Pada Ibu Hamilgia amandaAinda não há avaliações
- Publishers Take NoteaxicvDocumento1 páginaPublishers Take Noteaxicvwoolonion5Ainda não há avaliações
- Neonatal Sepsis 2018Documento40 páginasNeonatal Sepsis 2018Abraham AnaelyAinda não há avaliações
- Cytokines and Prostaglandins in Immune Homeostasis and Tissue Destruction in Periodontal DiseaseDocumento32 páginasCytokines and Prostaglandins in Immune Homeostasis and Tissue Destruction in Periodontal DiseaseDiana GomezAinda não há avaliações
- Neonatal Sepsis in The Very Low Birth Weight Preterm Infants: Part 1: Review of Patho-PhysiologyDocumento10 páginasNeonatal Sepsis in The Very Low Birth Weight Preterm Infants: Part 1: Review of Patho-PhysiologyjukunkAinda não há avaliações
- Active Immunization Long DOCPDFDocumento31 páginasActive Immunization Long DOCPDFEnder IzaguirreAinda não há avaliações
- Inmunidad y SepsisDocumento6 páginasInmunidad y SepsisAmmarie SanchezAinda não há avaliações
- Cellular Immune Responses in The Pathophysiology of 2021Documento24 páginasCellular Immune Responses in The Pathophysiology of 2021Zakkiyatus ZainiyahAinda não há avaliações
- Role of Immunoglobulins in Neonatal SepsisDocumento6 páginasRole of Immunoglobulins in Neonatal SepsisMuhar RandiAinda não há avaliações
- Christ The King College College of Nursing and IHAP Calbayog CityDocumento4 páginasChrist The King College College of Nursing and IHAP Calbayog CityMannuelle GacudAinda não há avaliações
- Diagnostics 12 00453Documento15 páginasDiagnostics 12 00453Fabiola Vania FeliciaAinda não há avaliações
- Jurnal BPHDocumento15 páginasJurnal BPHdewasayogaAinda não há avaliações
- Immune System - 10pDocumento10 páginasImmune System - 10pLinh LeAinda não há avaliações
- Fast Facts: Managing Immune-Related Adverse Events in OncologyNo EverandFast Facts: Managing Immune-Related Adverse Events in OncologyAinda não há avaliações
- A Comprehensive Guide To Managing AutismDocumento89 páginasA Comprehensive Guide To Managing Autismapi-3696876100% (4)
- Infancy To Old Age Immune SysytemDocumento9 páginasInfancy To Old Age Immune Sysytemlinnet17Ainda não há avaliações
- Type 1 Diabetes Mellitus POSTERDocumento1 páginaType 1 Diabetes Mellitus POSTERLeandro FigueiraAinda não há avaliações
- Immunology of Pregnancy1Documento50 páginasImmunology of Pregnancy1Ekiran BabajideAinda não há avaliações
- Basic ImmunologyDocumento12 páginasBasic Immunologyjony_phurailatpamAinda não há avaliações