Você também pode gostar
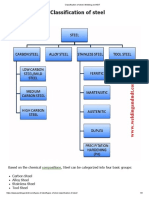
- Classification of Steel - Welding and NDTDocumento3 páginasClassification of Steel - Welding and NDTAshif Iqubal100% (1)
- Ethem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)Documento209 páginasEthem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)riddhiAinda não há avaliações
- Wa0044 PDFDocumento337 páginasWa0044 PDFYamilAinda não há avaliações
- Palm Based Non Hydrogenated Creamer PDFDocumento4 páginasPalm Based Non Hydrogenated Creamer PDFbellesuperAinda não há avaliações
- ENZYME KINETICSDocumento11 páginasENZYME KINETICSDianne Villanueva100% (1)
- Column and Thin Layer ChromatographyDocumento5 páginasColumn and Thin Layer Chromatographymarilujane80% (5)
- Ubiquinone Biosynthesis of QuinoneDocumento12 páginasUbiquinone Biosynthesis of QuinoneFrancisco EspinosaAinda não há avaliações
- Gene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialDocumento6 páginasGene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialHuong TranAinda não há avaliações
- CoQ10-Biosynthesis-1-s2.0-S0005272816300573-mainDocumento6 páginasCoQ10-Biosynthesis-1-s2.0-S0005272816300573-maingjlynxAinda não há avaliações
- Anthocyanin Synthesis in Maize Aleurone Tissue: Conek@missouri - EduDocumento2 páginasAnthocyanin Synthesis in Maize Aleurone Tissue: Conek@missouri - EduMonica Cabezas DipazAinda não há avaliações
- Research Project: Prof. Dr. Rehab HamedDocumento13 páginasResearch Project: Prof. Dr. Rehab HamedHassaninElaradyAinda não há avaliações
- Biotin Synthesis in Higher Plants: Purification and Characterization ofDocumento5 páginasBiotin Synthesis in Higher Plants: Purification and Characterization ofmyfreepaysite321Ainda não há avaliações
- Insights Into Phycoerythrobilin Biosynthesis Point Toward Metabolic ChannelingDocumento9 páginasInsights Into Phycoerythrobilin Biosynthesis Point Toward Metabolic ChannelingAvani KaushalAinda não há avaliações
- Chloroplast Import of Four Carotenoid Biosynthetic EnzymesDocumento9 páginasChloroplast Import of Four Carotenoid Biosynthetic EnzymesNithin RAinda não há avaliações
- Seminario 1-2022-1Documento13 páginasSeminario 1-2022-1Elias Gerardo Pardo LlamoccaAinda não há avaliações
- Silva Et Al. 2021Documento11 páginasSilva Et Al. 2021Davi OliveiraAinda não há avaliações
- Structural and Biological Aspects of Carotenoid CleavageDocumento13 páginasStructural and Biological Aspects of Carotenoid CleavageMinh Chau NguyenAinda não há avaliações
- Alkaloids and OthersDocumento24 páginasAlkaloids and OtherszakybaaAinda não há avaliações
- Bacterial Phospholipase A: Structure and Function of An Integral Membrane PhospholipaseDocumento11 páginasBacterial Phospholipase A: Structure and Function of An Integral Membrane PhospholipaseDiana SahoneroAinda não há avaliações
- Botryococcus Braunii, Race B: Isolation and Characterization of Two Squalene Epoxidase Genes FromDocumento14 páginasBotryococcus Braunii, Race B: Isolation and Characterization of Two Squalene Epoxidase Genes FromCarlos A. PonceeAinda não há avaliações
- Redox InfoDocumento11 páginasRedox InfoCasandra Valencia OjedaAinda não há avaliações
- Mitochondrial Diseases and Genetic Defects of ATP SynthaseDocumento6 páginasMitochondrial Diseases and Genetic Defects of ATP SynthasewlymjzdwAinda não há avaliações
- (Department of Zoology,: P-ApocarotenalsDocumento1 página(Department of Zoology,: P-ApocarotenalsKrrliveAinda não há avaliações
- Phosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalDocumento12 páginasPhosphorylation of Enoyl-Acyl Carrier Protein Reductase Inha Impacts Mycobacterial Growth and SurvivalNini FarmAinda não há avaliações
- Metabolic Engineering of Cyanobacteria For Ethanol ProductionDocumento8 páginasMetabolic Engineering of Cyanobacteria For Ethanol ProductionPatrick FuAinda não há avaliações
- міоінозитолDocumento10 páginasміоінозитолDaryna DarynaAinda não há avaliações
- Redox BiologyDocumento13 páginasRedox BiologyLin LIAinda não há avaliações
- ATP Synthesis and Storage: Original ArticleDocumento15 páginasATP Synthesis and Storage: Original ArticleRiley RilanAinda não há avaliações
- Association Between Genetic Variants in The Coenzyme Q Metabolism and Coenzyme Q Status in HumansDocumento7 páginasAssociation Between Genetic Variants in The Coenzyme Q Metabolism and Coenzyme Q Status in HumansLUCIENE CRISTALDO ALBUQUERQUEAinda não há avaliações
- Hong 2005Documento11 páginasHong 2005tea.zakicAinda não há avaliações
- Enzymatic Total Synthesis of Gibberellin A4Documento8 páginasEnzymatic Total Synthesis of Gibberellin A4ranukahewageAinda não há avaliações
- Vitamin K Biosynthesis: A. MenaquinoneDocumento4 páginasVitamin K Biosynthesis: A. MenaquinoneArslan Muhammad EjazAinda não há avaliações
- Chemistry of Protein Kinases and Dephosphorylases: Advanced ArticleDocumento8 páginasChemistry of Protein Kinases and Dephosphorylases: Advanced ArticleazzaassAinda não há avaliações
- Ajikumar 2010Documento6 páginasAjikumar 2010Jonathan DiasAinda não há avaliações
- Topic 3.2Documento33 páginasTopic 3.2Salna Susan AbrahamAinda não há avaliações
- Þ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCDocumento12 páginasÞ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCgauravAinda não há avaliações
- Ousalem Et Al 2023Documento46 páginasOusalem Et Al 2023pampetraAinda não há avaliações
- High-Level Production of Recombinant Arenicola Marina Globin Chains in Escherichia Coli: A New Generation of Blood SubstituteDocumento12 páginasHigh-Level Production of Recombinant Arenicola Marina Globin Chains in Escherichia Coli: A New Generation of Blood SubstituteIstván PortörőAinda não há avaliações
- Self-Assembled Protein-Enzyme Nanoflower-Based Fluorescent Sensing For Protein BiomarkerDocumento8 páginasSelf-Assembled Protein-Enzyme Nanoflower-Based Fluorescent Sensing For Protein BiomarkerAlvaro Matus CAinda não há avaliações
- Manipulation of Malic Enzyme in Saccharomyces Cerevisiae For Increasing NADPH ProductionDocumento12 páginasManipulation of Malic Enzyme in Saccharomyces Cerevisiae For Increasing NADPH ProductionJesús Miguel Jacinto NavaAinda não há avaliações
- COQ7 phenotipic, molecular functioningDocumento13 páginasCOQ7 phenotipic, molecular functioningLaura A M MAinda não há avaliações
- FotosintesisDocumento6 páginasFotosintesisNorma YantiAinda não há avaliações
- Fotosintesis PDFDocumento6 páginasFotosintesis PDFNorma YantiAinda não há avaliações
- Artículo RehabilitaciónDocumento10 páginasArtículo RehabilitaciónWilliam rodriguez ramosAinda não há avaliações
- Bacterial Phenylalanine and Phenylacetate Catabolic Pathway RevealedDocumento6 páginasBacterial Phenylalanine and Phenylacetate Catabolic Pathway RevealedKarla KetiAinda não há avaliações
- Inorganic Polyphosphates in Mitochondria Review 2010Documento7 páginasInorganic Polyphosphates in Mitochondria Review 2010Baranov MaximAinda não há avaliações
- Figuras Insight Into The Evolution of Microbial Metabolism From The Deep-Branching Bacterium, Thermovibrio AmmonificansDocumento14 páginasFiguras Insight Into The Evolution of Microbial Metabolism From The Deep-Branching Bacterium, Thermovibrio AmmonificansIara GAinda não há avaliações
- Biochemical and Biophysical Research CommunicationsDocumento5 páginasBiochemical and Biophysical Research CommunicationsFernando P. Molina-HerediaAinda não há avaliações
- Caracterización de La Olivetol Sintasa, Una Policétido Sintasa Implicada Supuestamente en La Ruta Biosintética de Los CannabinoidesDocumento6 páginasCaracterización de La Olivetol Sintasa, Una Policétido Sintasa Implicada Supuestamente en La Ruta Biosintética de Los CannabinoidesAnonymous 9oH7eQ2TAinda não há avaliações
- First Report: Yersinia Enterocolitica Recovered From Canine TonsilsDocumento5 páginasFirst Report: Yersinia Enterocolitica Recovered From Canine TonsilsMarcela TapiasAinda não há avaliações
- Babesia Canis Rossi: Characterization and Molecular Cloning of An Adenosine Kinase FromDocumento7 páginasBabesia Canis Rossi: Characterization and Molecular Cloning of An Adenosine Kinase FromSoare MarianAinda não há avaliações
- TMP 2 FEDocumento2 páginasTMP 2 FEFrontiersAinda não há avaliações
- Peroxisomal β-oxidation-A metabolic pathway with multiple functionsDocumento14 páginasPeroxisomal β-oxidation-A metabolic pathway with multiple functionsRoy WilsonAinda não há avaliações
- 5 Complementation of Apolipoprotein B mRNA Editing by Human Liver Accompanied by Secretion of Apolipoprotein B48 J. Biol. Chem.-1994-Giannoni-5932-6Documento5 páginas5 Complementation of Apolipoprotein B mRNA Editing by Human Liver Accompanied by Secretion of Apolipoprotein B48 J. Biol. Chem.-1994-Giannoni-5932-6Federico GiannoniAinda não há avaliações
- Cloning and expressing cellulase gene in silkworm using baculovirus systemDocumento8 páginasCloning and expressing cellulase gene in silkworm using baculovirus systemBhaskar RoyAinda não há avaliações
- Unesco - Eolss Sample Chapters: Bioplastic and Biopolymer ProductionDocumento10 páginasUnesco - Eolss Sample Chapters: Bioplastic and Biopolymer ProductionAliAliAinda não há avaliações
- Bioorganic & Medicinal ChemistryDocumento6 páginasBioorganic & Medicinal ChemistryRuthaiwan KongcharoenAinda não há avaliações
- Microbial Production of Biodegradable Polymers and Their Role in Cardiac Stent DevelopmentDocumento11 páginasMicrobial Production of Biodegradable Polymers and Their Role in Cardiac Stent Development2begeniusAinda não há avaliações
- Asembly VLDLDocumento4 páginasAsembly VLDLCornel GheraseAinda não há avaliações
- Art:10 1007/BF00173913Documento8 páginasArt:10 1007/BF00173913Leonardo SakamotoAinda não há avaliações
- 1 s2.0 0014579394009325 MainDocumento5 páginas1 s2.0 0014579394009325 MainMuhammad Andika PerdanaAinda não há avaliações
- 1 s2.0 S0005273614002454 MainDocumento8 páginas1 s2.0 S0005273614002454 MainAlex WasabiAinda não há avaliações
- A Minimal System Allowing Tubulation With Molecular Motors Pulling On Giant LiposomesDocumento6 páginasA Minimal System Allowing Tubulation With Molecular Motors Pulling On Giant LiposomesAbhishek KansalAinda não há avaliações
- Purine Metabolism PathwaysDocumento29 páginasPurine Metabolism Pathwaystrinitysugumar0% (1)
- Recent Advances in Polyphenol ResearchNo EverandRecent Advances in Polyphenol ResearchKumi YoshidaAinda não há avaliações
- Molecules: ISSN 1420-3049Documento25 páginasMolecules: ISSN 1420-3049Jesus LirioAinda não há avaliações
- Ceramic AsDocumento2 páginasCeramic AsJesus LirioAinda não há avaliações
- Solanum Mammosum LDocumento21 páginasSolanum Mammosum LJesus LirioAinda não há avaliações
- Antibacterial Activity of Tannins From The Leaves of Solanum Trilobatum Linn - Feb09dos PDFDocumento3 páginasAntibacterial Activity of Tannins From The Leaves of Solanum Trilobatum Linn - Feb09dos PDFarca_77@Ainda não há avaliações
- Transfer en CIA de Electrones en BacteriasDocumento6 páginasTransfer en CIA de Electrones en BacteriasJesus LirioAinda não há avaliações
- LOVely EnzymesDocumento10 páginasLOVely EnzymesJesus LirioAinda não há avaliações
- Brittany L. Hayes - Recent Advances in Microwave - Assisted SynthesisDocumento11 páginasBrittany L. Hayes - Recent Advances in Microwave - Assisted SynthesisnnnnjwAinda não há avaliações
- Plastics: Name: Taaha Muzaffar Imam ROLL NO.: 19011AA002 Sem/Sec: Ii/A Branch: B.Arch College: Spa' JnafauDocumento5 páginasPlastics: Name: Taaha Muzaffar Imam ROLL NO.: 19011AA002 Sem/Sec: Ii/A Branch: B.Arch College: Spa' JnafauTaaha Muzaffar ImamAinda não há avaliações
- Rambutan SunscreenDocumento7 páginasRambutan SunscreenAndrea Alvarado RoAinda não há avaliações
- Puresilk Salt ChlorinatorDocumento10 páginasPuresilk Salt Chlorinatornike_y2kAinda não há avaliações
- Experiment 2: Brinell Hardness TestDocumento5 páginasExperiment 2: Brinell Hardness TestseifAinda não há avaliações
- 1 ElectrochemistryExercise PDFDocumento46 páginas1 ElectrochemistryExercise PDFDivyanshi TiwaryAinda não há avaliações
- Prepared by Miss Unnati .M. Patel F.Y.M Pharm (Pqa) by DR A.D.Kulkerni Mpharm (Pharmaceutics)Documento29 páginasPrepared by Miss Unnati .M. Patel F.Y.M Pharm (Pqa) by DR A.D.Kulkerni Mpharm (Pharmaceutics)Fatima VessaliusAinda não há avaliações
- OVERVIEW (4 Points) : CH116 General and Organic Principles LabDocumento4 páginasOVERVIEW (4 Points) : CH116 General and Organic Principles Labapi-557329548Ainda não há avaliações
- Company Profile 4 April - CompressedDocumento10 páginasCompany Profile 4 April - CompressedPhilip PhilipsAinda não há avaliações
- Glyphosate Goker MSDS 1Documento7 páginasGlyphosate Goker MSDS 1Bima SitorusAinda não há avaliações
- Moisture RemovalDocumento9 páginasMoisture RemovalRafi AkbarAinda não há avaliações
- Content of The Dossier For Chemical Purity and Microbiological QualityDocumento23 páginasContent of The Dossier For Chemical Purity and Microbiological QualityjdemelloAinda não há avaliações
- Solid Hollow Plug PDFDocumento3 páginasSolid Hollow Plug PDFSandra DevannyAinda não há avaliações
- Chap 8 Ques - AnsDocumento11 páginasChap 8 Ques - AnsHaley WillhelmAinda não há avaliações
- Certificado de Calidad Caps SCH-40Documento1 páginaCertificado de Calidad Caps SCH-40Jesus CondoriAinda não há avaliações
- BooCax Disinfection Solution For Beijing 2022 Winter OlympicsDocumento21 páginasBooCax Disinfection Solution For Beijing 2022 Winter OlympicsRobotics BoocaxAinda não há avaliações
- GATE-Architecture Sample QuestionsDocumento6 páginasGATE-Architecture Sample QuestionsCharan ReddyAinda não há avaliações
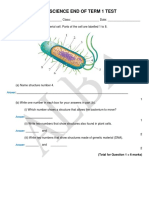
- Year 8 Science End of Term 1 Test: AnswerDocumento11 páginasYear 8 Science End of Term 1 Test: Answerchan myaeAinda não há avaliações
- 1 s2.0 S1687157X17300288 MainDocumento11 páginas1 s2.0 S1687157X17300288 Mainnagi jainAinda não há avaliações
- Tandem Dyes For Flow Cytometry, Quality Concerns, Beckman CoDocumento2 páginasTandem Dyes For Flow Cytometry, Quality Concerns, Beckman CocandiddreamsAinda não há avaliações
- Fmi Unit 2Documento86 páginasFmi Unit 2Pranav vigneshAinda não há avaliações
- Human Respiratory System Based On Law of ThermodynamicsDocumento9 páginasHuman Respiratory System Based On Law of ThermodynamicsfatimahAinda não há avaliações
- Mass-Transfer Diffusion Coefficients in Binary Systems: AppendixDocumento3 páginasMass-Transfer Diffusion Coefficients in Binary Systems: AppendixAnisaAinda não há avaliações
- Ekatalog 2023 Sulsel RajawaliDocumento50 páginasEkatalog 2023 Sulsel RajawaliSafria HamzaAinda não há avaliações