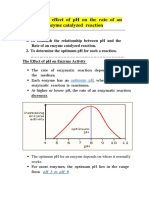
Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- The Effect of Temperature On The Hatching Success of Brine ShrimpDocumento2 páginasThe Effect of Temperature On The Hatching Success of Brine ShrimptahamidAinda não há avaliações
- The Influence of Giberelin On Stem GrowthDocumento11 páginasThe Influence of Giberelin On Stem GrowthDevina AlifahAinda não há avaliações
- General Biology - Chapter IDocumento10 páginasGeneral Biology - Chapter IG.k. Vinnan Rao100% (2)
- Effect of PH On Enzyme ActivityDocumento12 páginasEffect of PH On Enzyme ActivityAb AbAinda não há avaliações
- Dihybrid ProblemsDocumento6 páginasDihybrid ProblemsVicky GuzmánAinda não há avaliações
- Protein Synthesis Notes Sheet 20-21Documento4 páginasProtein Synthesis Notes Sheet 20-21Cassandra KankamAinda não há avaliações
- Blue BrainDocumento20 páginasBlue BrainAnkitaPatel009Ainda não há avaliações
- Immune Response in The Human BodyDocumento30 páginasImmune Response in The Human BodyjohnosborneAinda não há avaliações
- Menigeal LymphoidDocumento10 páginasMenigeal LymphoidRenju KuriakoseAinda não há avaliações
- MotivationDocumento2 páginasMotivationPrachi Srivastava100% (1)
- PGA102 Presentation 1 (Eng. Ver.)Documento18 páginasPGA102 Presentation 1 (Eng. Ver.)Lee Wai KeatAinda não há avaliações
- Paired Passages - PresentationDocumento17 páginasPaired Passages - Presentationsuchi ravaliaAinda não há avaliações
- Eye - MCQs CorrectDocumento207 páginasEye - MCQs CorrectMaaz KhanAinda não há avaliações
- Activity No. 7.1 BloodDocumento2 páginasActivity No. 7.1 BloodDree SermanAinda não há avaliações
- Beckers World of The Cell 9th Edition Hardin Test Bank DownloadDocumento23 páginasBeckers World of The Cell 9th Edition Hardin Test Bank Downloaddaddockstudderyxeq100% (27)
- Guyabano Plant ResearchDocumento2 páginasGuyabano Plant ResearchAnnie OmlangAinda não há avaliações
- (English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)Documento78 páginas(English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)acromontiAinda não há avaliações
- 1 Human Anatomy Physiology With PathologyDocumento41 páginas1 Human Anatomy Physiology With PathologyEd dela PenaAinda não há avaliações
- Pathophysiology of Conjoined TwinsDocumento1 páginaPathophysiology of Conjoined TwinsPrincess Loreto AlegreAinda não há avaliações
- Summative Assessment QuizDocumento3 páginasSummative Assessment Quizapi-379149339Ainda não há avaliações
- Minimal Intervention Dentistry in General Dentistry PDFDocumento10 páginasMinimal Intervention Dentistry in General Dentistry PDFmostafahassan777Ainda não há avaliações
- Reference RangesDocumento8 páginasReference RangesKru PrimeAinda não há avaliações
- Dolly The SheepDocumento7 páginasDolly The SheepRhan AlcantaraAinda não há avaliações
- Car - Structural Bases For Shaping of DolinesDocumento18 páginasCar - Structural Bases For Shaping of DolinesCaegeoAinda não há avaliações
- 6 The Main PeriodsDocumento5 páginas6 The Main PeriodsUtami MAinda não há avaliações
- Arndt Schmidt-Denter Auer Weitere 2003Documento15 páginasArndt Schmidt-Denter Auer Weitere 2003Louis Fetilo FabunanAinda não há avaliações
- Immobilization of Catalase Via Adsorption Into NatDocumento6 páginasImmobilization of Catalase Via Adsorption Into NatErwin Guillermo Valdizón WinterAinda não há avaliações
- Lecture 3 Cell Cycle - QuestionsDocumento6 páginasLecture 3 Cell Cycle - Questions中华雅思王Ainda não há avaliações
- Dalandan (Citrus Aurantium Linn.) Peelings and Calamansi (Citrofortunella Microcarpa) Extract As An Alternative Fire Ant KillerDocumento6 páginasDalandan (Citrus Aurantium Linn.) Peelings and Calamansi (Citrofortunella Microcarpa) Extract As An Alternative Fire Ant KillerMaryknoll BernabeAinda não há avaliações
- 088 11MS9419 Print PDFDocumento11 páginas088 11MS9419 Print PDFNehaAinda não há avaliações