Você também pode gostar
- Position Paper-Special TopicsDocumento20 páginasPosition Paper-Special TopicsGabriela FernandezAinda não há avaliações
- 2018 Ei-2Documento8 páginas2018 Ei-2MichelleAinda não há avaliações
- Artigo 1Documento3 páginasArtigo 1Atarissis DiasAinda não há avaliações
- Accepted Manuscript: 10.1016/j.gecco.2018.e00412Documento23 páginasAccepted Manuscript: 10.1016/j.gecco.2018.e00412ArlittAmyLozanoPovisAinda não há avaliações
- NameDocumento9 páginasNamejosephmainam9Ainda não há avaliações
- Uncertainty and Climate Change: 1. OverviewDocumento37 páginasUncertainty and Climate Change: 1. OverviewCaroAinda não há avaliações
- Topic 1 IB GuideDocumento6 páginasTopic 1 IB GuideMichael SmithAinda não há avaliações
- Tipe 02Documento20 páginasTipe 02Nanda desima silalahiAinda não há avaliações
- Bookshelf: Search Termsearch DatabaseDocumento31 páginasBookshelf: Search Termsearch DatabaseMeliza Qquelcca MamanquiAinda não há avaliações
- Scientific Uncertainties: Unscientific ArgumentsDocumento8 páginasScientific Uncertainties: Unscientific ArgumentsbillpaparounisAinda não há avaliações
- International Journal of Computational Engineering Research (IJCER)Documento22 páginasInternational Journal of Computational Engineering Research (IJCER)International Journal of computational Engineering research (IJCER)Ainda não há avaliações
- Climate Change AdaptationDocumento118 páginasClimate Change AdaptationopulitheAinda não há avaliações
- Annotated Bibliography - Essay 2 CorrectionDocumento7 páginasAnnotated Bibliography - Essay 2 Correctionapi-644176765Ainda não há avaliações
- The Article Talks About How Costa Rica: P3.1 Argument and AnalysisDocumento1 páginaThe Article Talks About How Costa Rica: P3.1 Argument and Analysisapi-699145220Ainda não há avaliações
- Extreme Climatic EventsDocumento8 páginasExtreme Climatic EventsAlexa MariusAinda não há avaliações
- Ecological Laws For Agroecological Design: The Need For More Organized Collaboration in Producing, Evaluating and Updating Ecological GeneralizationsDocumento20 páginasEcological Laws For Agroecological Design: The Need For More Organized Collaboration in Producing, Evaluating and Updating Ecological GeneralizationsRicardinho1987Ainda não há avaliações
- Naeem Et Al 2012 Biodiversity ExtinctionsDocumento7 páginasNaeem Et Al 2012 Biodiversity ExtinctionsGloria AcostaAinda não há avaliações
- Does Ecology Need A Technical Revolution - Honours Course Extinction - Sebastiaan de Jager 11301430Documento5 páginasDoes Ecology Need A Technical Revolution - Honours Course Extinction - Sebastiaan de Jager 11301430api-408361250Ainda não há avaliações
- Climate Change PDFDocumento230 páginasClimate Change PDFTarun MattaparthyAinda não há avaliações
- Research Paper of Climate ChangeDocumento10 páginasResearch Paper of Climate Changeefj02jba100% (1)
- 8 Data Gaps, Uncertainties and Future Needs: Need To Address UncertaintyDocumento15 páginas8 Data Gaps, Uncertainties and Future Needs: Need To Address UncertaintyCIO-CIOAinda não há avaliações
- Resilience and Sustainable DevelopmentDocumento11 páginasResilience and Sustainable DevelopmentMathiyazhagan KAinda não há avaliações
- Attribution ExtreAttribution of Extreme Weather Events in The Context of Climate Changeme Weather Brief FinalDocumento4 páginasAttribution ExtreAttribution of Extreme Weather Events in The Context of Climate Changeme Weather Brief FinalNational Academies of Science, Engineering, and MedicineAinda não há avaliações
- Melbinger 2015Documento11 páginasMelbinger 2015yalocim666Ainda não há avaliações
- Impacts of Climate Change On BiodiversityDocumento4 páginasImpacts of Climate Change On BiodiversityNimo MohamudAinda não há avaliações
- Hooper Et Al 2005Documento33 páginasHooper Et Al 2005Jhonatan Gutierrez100% (1)
- Population Approach in Ecotoxicology (Stress Ecology)Documento12 páginasPopulation Approach in Ecotoxicology (Stress Ecology)3STGameAinda não há avaliações
- Waldvogel Et Al-2020-Evolution LettersDocumento15 páginasWaldvogel Et Al-2020-Evolution LettersAdhst GuhrzisAinda não há avaliações
- Causality by David CoxDocumento21 páginasCausality by David CoxQianhan LinAinda não há avaliações
- Environment and Happiness Valuation of Air Pollution Using Life Satisfaction DataDocumento13 páginasEnvironment and Happiness Valuation of Air Pollution Using Life Satisfaction DataMilan VeljkovicAinda não há avaliações
- Complete Research Paper About Climate ChangeDocumento6 páginasComplete Research Paper About Climate Changegz8qs4dn100% (1)
- Assessing The Impact of Climate Change On Biodiversity Hotspots A Comparative StudyDocumento3 páginasAssessing The Impact of Climate Change On Biodiversity Hotspots A Comparative Studymisaj17879Ainda não há avaliações
- English ResearchDocumento4 páginasEnglish ResearchHAHAHAAinda não há avaliações
- climatechangechap3WHO Intnl ConcesusDocumento18 páginasclimatechangechap3WHO Intnl ConcesusrevaAinda não há avaliações
- Annotated BibliographyDocumento5 páginasAnnotated Bibliographysiaf khanAinda não há avaliações
- ClimatologyDocumento3 páginasClimatologyfredtash96Ainda não há avaliações
- Literature Review Climate Change Vulnerability Assessment Risk Assessment and Adaptation ApproachesDocumento5 páginasLiterature Review Climate Change Vulnerability Assessment Risk Assessment and Adaptation ApproachesfgwuwoukgAinda não há avaliações
- Kareiva, P. and Fuller - Beyond Resilience - How To Better Prepare For The Pro - Found Disruption of The AnthropoceneDocumento27 páginasKareiva, P. and Fuller - Beyond Resilience - How To Better Prepare For The Pro - Found Disruption of The AnthropoceneAriel BärAinda não há avaliações
- Research Paper Using CorrelationDocumento6 páginasResearch Paper Using Correlationafnhekkghifrbm100% (1)
- Group D7 BIO 31.02 Project ProposalDocumento1 páginaGroup D7 BIO 31.02 Project ProposalPaul PanizaAinda não há avaliações
- GFS - Tipping Points - Main ReportDocumento32 páginasGFS - Tipping Points - Main ReportModi JesusAinda não há avaliações
- An Introduction To Epidemiologic and Statistical Methods Useful in Environmental EpidemiologyDocumento8 páginasAn Introduction To Epidemiologic and Statistical Methods Useful in Environmental EpidemiologyAndi Fahira NurAinda não há avaliações
- Document 1Documento21 páginasDocument 1jamaica.apasAinda não há avaliações
- Berk & Fovell 1998Documento43 páginasBerk & Fovell 1998ktiiti60Ainda não há avaliações
- Tiff Ja Environmental ScienceDocumento3 páginasTiff Ja Environmental SciencekyanglisondraAinda não há avaliações
- Article SummaryDocumento6 páginasArticle Summarybrian ngugiAinda não há avaliações
- Con Secuencia SDocumento9 páginasCon Secuencia Sdatura49Ainda não há avaliações
- Correlation of Environmental Factors On The Spread of Microbial Infections Disease in The PhilippinesDocumento6 páginasCorrelation of Environmental Factors On The Spread of Microbial Infections Disease in The PhilippinesAbi NeriAinda não há avaliações
- PHD Thesis Climate Change AdaptationDocumento7 páginasPHD Thesis Climate Change Adaptationstaceycruzwashington100% (2)
- Effect of Climate Change To The Number of Harvest of Selected Farmers in Barangay Lucsuhin, Alfonso, CaviteDocumento9 páginasEffect of Climate Change To The Number of Harvest of Selected Farmers in Barangay Lucsuhin, Alfonso, CaviteMISSKPOP tvAinda não há avaliações
- Calentamiento GlobalDocumento22 páginasCalentamiento GlobalAnonymous 2DvIUvkCAinda não há avaliações
- Climate: Livestock Under Climate Change: A Systematic Review of Impacts and AdaptationDocumento17 páginasClimate: Livestock Under Climate Change: A Systematic Review of Impacts and AdaptationDept. of AH Economics MVCAinda não há avaliações
- Zhai2018 Article AReviewOfClimateChangeAttributDocumento22 páginasZhai2018 Article AReviewOfClimateChangeAttributfaewfefaAinda não há avaliações
- Tugas Prom Gizi12Documento8 páginasTugas Prom Gizi12Krisna AlfianurAinda não há avaliações
- Recent Evidence For The Climate Change Threat To Lepidoptera and Other InsectsDocumento10 páginasRecent Evidence For The Climate Change Threat To Lepidoptera and Other InsectsBreno LoboAinda não há avaliações
- Scientists' Warning On AffluenceDocumento10 páginasScientists' Warning On AffluenceÉditions DédicacesAinda não há avaliações
- VATER AssociationDocumento2 páginasVATER AssociationAreef MuarifAinda não há avaliações
- Cri Du Chat SyndromeDocumento2 páginasCri Du Chat SyndromeAreef MuarifAinda não há avaliações
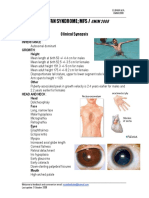
- Sotos Syndrome / Cerebral Gigantism /: Clinical SynopsisDocumento2 páginasSotos Syndrome / Cerebral Gigantism /: Clinical SynopsisAreef MuarifAinda não há avaliações
- Alagille SyndromeDocumento2 páginasAlagille SyndromeAreef MuarifAinda não há avaliações
- Silver Rassell SyndromeDocumento2 páginasSilver Rassell SyndromeAreef MuarifAinda não há avaliações
- Progeria SyndromeDocumento1 páginaProgeria SyndromeAreef MuarifAinda não há avaliações
- Rett Syndrome RTT /: OMIM2008 Clinical Synopsis Clinical Synopsis Clinical Synopsis Clinical SynopsisDocumento2 páginasRett Syndrome RTT /: OMIM2008 Clinical Synopsis Clinical Synopsis Clinical Synopsis Clinical SynopsisAreef MuarifAinda não há avaliações
- Noonan SyndromeDocumento3 páginasNoonan SyndromeAreef MuarifAinda não há avaliações
- Fanconi AnemiaDocumento2 páginasFanconi AnemiaAreef MuarifAinda não há avaliações
- Marfan SyndromeDocumento3 páginasMarfan SyndromeAreef MuarifAinda não há avaliações
- FragileX SyndromeDocumento2 páginasFragileX SyndromeAreef MuarifAinda não há avaliações
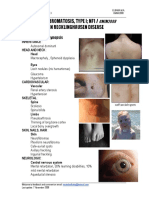
- NeurofibromatosisType IDocumento2 páginasNeurofibromatosisType IAreef MuarifAinda não há avaliações
- CHARGE AssociationDocumento2 páginasCHARGE AssociationAreef MuarifAinda não há avaliações
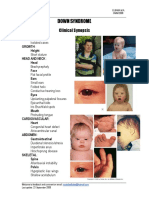
- Down SyndromeDocumento2 páginasDown SyndromeAreef MuarifAinda não há avaliações
- Neurodevelopment CMV PDFDocumento8 páginasNeurodevelopment CMV PDFAreef MuarifAinda não há avaliações
- Manajemen Hearing Loss PDFDocumento15 páginasManajemen Hearing Loss PDFAreef MuarifAinda não há avaliações
- Cunha Et Al., 2019 - Art 2Documento9 páginasCunha Et Al., 2019 - Art 2Erika SierraAinda não há avaliações
- DiGeorge SyndromeDocumento3 páginasDiGeorge SyndromeAreef MuarifAinda não há avaliações
- Angelman Syndrome As Angelman Syndrome As Angelman Syndrome As Angelman Syndrome AsDocumento2 páginasAngelman Syndrome As Angelman Syndrome As Angelman Syndrome As Angelman Syndrome AsAreef MuarifAinda não há avaliações
- Journal 1 PDFDocumento9 páginasJournal 1 PDFAntónio Ayrton WidiastaraAinda não há avaliações
- gjmbsv3n5 16Documento6 páginasgjmbsv3n5 16Areef MuarifAinda não há avaliações
- Pediatric CardiologyDocumento6 páginasPediatric CardiologyAreef MuarifAinda não há avaliações
- Offline - POSBINDU Meri 10Documento21 páginasOffline - POSBINDU Meri 10Areef MuarifAinda não há avaliações
- DikoDocumento7 páginasDikoAreef MuarifAinda não há avaliações
- Hiperbilirubinemia by Ucsf HospitalDocumento3 páginasHiperbilirubinemia by Ucsf HospitalGalenica HoneyAinda não há avaliações
- Herbal Medicine: Current Status and The FutureDocumento8 páginasHerbal Medicine: Current Status and The FuturexiuhtlaltzinAinda não há avaliações
- Dede 5Documento5 páginasDede 5Areef MuarifAinda não há avaliações
- Dede2 PretermDocumento9 páginasDede2 PretermAreef MuarifAinda não há avaliações
- NICE Guidelines PDFDocumento41 páginasNICE Guidelines PDFglruvalAinda não há avaliações
- 85120030402Documento3 páginas85120030402Areef MuarifAinda não há avaliações
- 000000000001016591Documento212 páginas000000000001016591chowhkAinda não há avaliações
- 2019 05 Exam SRM SyllabusDocumento5 páginas2019 05 Exam SRM SyllabusSujith GopinathanAinda não há avaliações
- 4 - Simple Linear Regression I 2022-23Documento25 páginas4 - Simple Linear Regression I 2022-23tedAinda não há avaliações
- Richard Carrier Proving History ReviewDocumento32 páginasRichard Carrier Proving History ReviewTim Hendrix100% (2)
- Forest Ecology and Management: A C B C C C D CDocumento9 páginasForest Ecology and Management: A C B C C C D CJanisson BatistaAinda não há avaliações
- EconometricsDocumento8 páginasEconometricssamidan tubeAinda não há avaliações
- Deep Learning - Fundamentals, Theory and Applications 2019 PDFDocumento168 páginasDeep Learning - Fundamentals, Theory and Applications 2019 PDFismake31100% (2)
- Article BiophInt Assessing Shelf Life From Real Time and Accelerated Studies 2003Documento7 páginasArticle BiophInt Assessing Shelf Life From Real Time and Accelerated Studies 2003Tom NuytsAinda não há avaliações
- Estimating Project S-Curves Using Polynomial Function and Neural Networks Chao, ChienDocumento9 páginasEstimating Project S-Curves Using Polynomial Function and Neural Networks Chao, ChienisenefretAinda não há avaliações
- Probabilistic Approach For Durability Design of Concrete Structures in Marine EnvironmentsDocumento9 páginasProbabilistic Approach For Durability Design of Concrete Structures in Marine EnvironmentsTanvirH.ChowdhuryAinda não há avaliações
- Research ParadigmsDocumento10 páginasResearch ParadigmsAroosa ZaheerAinda não há avaliações
- Postresp UmDocumento328 páginasPostresp UmGuilherme LealAinda não há avaliações
- QMM Epgdm 5Documento58 páginasQMM Epgdm 5manish guptaAinda não há avaliações
- Chap 5 MCQDocumento12 páginasChap 5 MCQShalabh TewariAinda não há avaliações
- Dissecting The Cycle: A Methodological Investigation: Don Harding, Adrian PaganDocumento17 páginasDissecting The Cycle: A Methodological Investigation: Don Harding, Adrian PaganJavier PeñaAinda não há avaliações
- Time Series Analysis - Univariate and Multivariate Methods by William Wei PDFDocumento634 páginasTime Series Analysis - Univariate and Multivariate Methods by William Wei PDFAnuj Sachdev100% (3)
- NLP and EntropyDocumento54 páginasNLP and EntropymyoxygenAinda não há avaliações
- Formulation and Evaluation of Transdermal Patch of Repaglinide PDFDocumento9 páginasFormulation and Evaluation of Transdermal Patch of Repaglinide PDFapa menAinda não há avaliações
- Air Quality Model Performance EvaluationDocumento30 páginasAir Quality Model Performance EvaluationStefania IordacheAinda não há avaliações
- Monte CarloDocumento59 páginasMonte Carloadiraju07Ainda não há avaliações
- Baysian Analysis NotesDocumento30 páginasBaysian Analysis NotesOmer ShafiqAinda não há avaliações
- 500 Data Science Interview Questions and Answers - Vamsee Puligadda PDFDocumento141 páginas500 Data Science Interview Questions and Answers - Vamsee Puligadda PDFFranco Abanto67% (6)
- Uncertainty Analysis in Ship Performance Monitoring Draft Under ReviewDocumento23 páginasUncertainty Analysis in Ship Performance Monitoring Draft Under Reviewasit_kgpAinda não há avaliações
- Statistics For Financial Engineering: Some R Examples: David RuppertDocumento60 páginasStatistics For Financial Engineering: Some R Examples: David RuppertgauravroongtaAinda não há avaliações
- Chapter 1Documento24 páginasChapter 1Pei PeiAinda não há avaliações
- ConceptScript 2.1 PDFDocumento4 páginasConceptScript 2.1 PDFPuji SnAinda não há avaliações
- Stochastic User EquilibriumDocumento18 páginasStochastic User EquilibriumTseganheAinda não há avaliações
- 05 Class RegressionCorrelationDocumento57 páginas05 Class RegressionCorrelationJipeeZedAinda não há avaliações
- SMS - QBDocumento5 páginasSMS - QBAtul Dhar dubeyAinda não há avaliações
- HLM7 PDFDocumento366 páginasHLM7 PDFAnonymous tcPTEUOGJAinda não há avaliações