Você também pode gostar
- MIT Lecture 6 5.07 Biochemistry LectureDocumento16 páginasMIT Lecture 6 5.07 Biochemistry LectureakiridoAinda não há avaliações
- Sample ProblemDocumento1 páginaSample ProblemfintastellaAinda não há avaliações
- Biosynthesis of HemoglobinDocumento53 páginasBiosynthesis of HemoglobinAsep FauziAinda não há avaliações
- Voet - Chapt - 12 Properties of EnzymesDocumento102 páginasVoet - Chapt - 12 Properties of Enzymestelmo flowAinda não há avaliações
- Glycolysis and TCA Cycle - ShuttlesDocumento1 páginaGlycolysis and TCA Cycle - ShuttlesDr. SHIVA AITHALAinda não há avaliações
- 5 Enzyme Kinetics-InhibitionDocumento40 páginas5 Enzyme Kinetics-InhibitionJoel SmolanoffAinda não há avaliações
- Elisa & RiaDocumento4 páginasElisa & Riadihajum3Ainda não há avaliações
- Liver FunctionDocumento30 páginasLiver FunctionAra ShirikaAinda não há avaliações
- What Is The Oral Glucose Tolerance TestDocumento2 páginasWhat Is The Oral Glucose Tolerance TestBianca Camille100% (1)
- SGOT ASAT Kit Reitman Frankel MethodDocumento3 páginasSGOT ASAT Kit Reitman Frankel MethodShribagla MukhiAinda não há avaliações
- Excel 2007 TutorialDocumento5 páginasExcel 2007 TutorialromelcarvajalAinda não há avaliações
- Liver Function Test: Prepared By: Siti Norhaiza Binti HadzirDocumento33 páginasLiver Function Test: Prepared By: Siti Norhaiza Binti Hadzirmhafiziab100% (1)
- Dyslipidemia - HyperlipoproteinemiaDocumento43 páginasDyslipidemia - HyperlipoproteinemiaDarien LiewAinda não há avaliações
- GluconeogenesisDocumento11 páginasGluconeogenesisMithilesh RautAinda não há avaliações
- Elisa: From A ZDocumento66 páginasElisa: From A ZUsman WaheedAinda não há avaliações
- Urine PreservativesDocumento14 páginasUrine PreservativesHemamalini Ramaseshan100% (2)
- 06.disorder of Carbohydrate MetabolismDocumento47 páginas06.disorder of Carbohydrate MetabolismRizka NizarAinda não há avaliações
- Protein TargettingDocumento11 páginasProtein TargettingPMIB Matrikulasi FKUI 2018/2019Ainda não há avaliações
- Antigen Antibody ReactionDocumento3 páginasAntigen Antibody ReactionSai Sridhar100% (1)
- Tandem Mass Spectrometry MSMSDocumento22 páginasTandem Mass Spectrometry MSMSFegalma Eot100% (1)
- Interferences in Protein Electrophoresis Mahajan SlidesDocumento21 páginasInterferences in Protein Electrophoresis Mahajan SlidesDarshana JuvekarAinda não há avaliações
- Elisa Reader & Microplate Washer: Farheen Saba PH.D Ist Year Deptt. of Zoology ManuuDocumento30 páginasElisa Reader & Microplate Washer: Farheen Saba PH.D Ist Year Deptt. of Zoology ManuuPious SabaAinda não há avaliações
- 4 - HemoglobinopathiesDocumento19 páginas4 - HemoglobinopathiesHamzehAinda não há avaliações
- CFTDocumento9 páginasCFTWad Kafouri100% (1)
- Automated Cell Counts HB Estimation MethodsDocumento10 páginasAutomated Cell Counts HB Estimation Methodsjohn mwambuAinda não há avaliações
- Electrothermal Industries PDFDocumento10 páginasElectrothermal Industries PDFAna Marie AllamAinda não há avaliações
- Immunohematology Handout PDFDocumento9 páginasImmunohematology Handout PDFTanveerAinda não há avaliações
- Radioimmunoassay (Ria) 3Documento21 páginasRadioimmunoassay (Ria) 3Sapna JainAinda não há avaliações
- Plasma ProteinsDocumento8 páginasPlasma ProteinsAaryan50% (2)
- Biosynthesis of HemoglobinDocumento42 páginasBiosynthesis of Hemoglobin95kodok85Ainda não há avaliações
- Plasma ProteinDocumento79 páginasPlasma ProteinnadieAinda não há avaliações
- Mechanism of Hormone ActionDocumento2 páginasMechanism of Hormone Actionegfr3yfgAinda não há avaliações
- Name: Kristine Joy Atos Block: BSN 1-D Practice ProblemsDocumento6 páginasName: Kristine Joy Atos Block: BSN 1-D Practice ProblemsJenz Hope Segui Novela100% (3)
- G8 B-Thall Chromatogram Interpretation Guide1Documento58 páginasG8 B-Thall Chromatogram Interpretation Guide1Kwok Hoi ShanAinda não há avaliações
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocumento72 páginasChapter 2: Diagnostic Enzymology: E+S Es E+PQananiisa Lammii Gammada100% (1)
- Elisa (Enzyme Linked Immunosorbant Assay)Documento4 páginasElisa (Enzyme Linked Immunosorbant Assay)Linette GuillermoAinda não há avaliações
- Urea Nitrogen AnalyzerDocumento16 páginasUrea Nitrogen AnalyzerSkywalker_92Ainda não há avaliações
- Biochemical Changes in PregnancyDocumento6 páginasBiochemical Changes in PregnancyDr.Aaisha Mohammed Al BalushiAinda não há avaliações
- Protein SynthesisDocumento21 páginasProtein Synthesisnorma thamrinAinda não há avaliações
- Enzymes - PPT 1Documento54 páginasEnzymes - PPT 1Cesar Augusto Airampo Macedo100% (1)
- Lecture 10 - Plasma Proteins - 18 Sep 2006Documento34 páginasLecture 10 - Plasma Proteins - 18 Sep 2006api-3703352100% (3)
- Cyanmethemoglobin Refers To A Relatively NonDocumento5 páginasCyanmethemoglobin Refers To A Relatively NonZahir Jayvee Gayak IIAinda não há avaliações
- MCQ MembranesDocumento10 páginasMCQ MembranesMarilyne RizkAinda não há avaliações
- Complement Fixation Test (CFT) : Microbiology PharmacyDocumento11 páginasComplement Fixation Test (CFT) : Microbiology PharmacyAisya Amalia MuslimaAinda não há avaliações
- Triple Sugar Iron AgarDocumento3 páginasTriple Sugar Iron AgarmaniAinda não há avaliações
- 2.plasma ProteinsDocumento5 páginas2.plasma ProteinsAnusuya SAinda não há avaliações
- URIC ACID LyphoDocumento2 páginasURIC ACID LyphoDharmesh Patel50% (2)
- Red Blood Cell Indices: DR KatekDocumento10 páginasRed Blood Cell Indices: DR KatekNathan Lupiya100% (1)
- NPN ReviewDocumento46 páginasNPN Reviewmika de guzmanAinda não há avaliações
- Albumin - BCG PDFDocumento2 páginasAlbumin - BCG PDFHamed Alsalhy100% (1)
- Estimation of AlbuminDocumento2 páginasEstimation of AlbuminAnand VeerananAinda não há avaliações
- Glucose Tolerance TestDocumento11 páginasGlucose Tolerance TestDharmikPatelAinda não há avaliações
- Kidney Function TestsDocumento50 páginasKidney Function TestsStephen Fritz OreAinda não há avaliações
- Mechanism of Enzyme ActionDocumento2 páginasMechanism of Enzyme ActionBlazy InhumangAinda não há avaliações
- Biochemistry NotesDocumento14 páginasBiochemistry NotesJocabed RadillaAinda não há avaliações
- Autoimmune DiseasesDocumento43 páginasAutoimmune DiseasesAhmed NajmAinda não há avaliações
- EnzymeKinetics PDFDocumento21 páginasEnzymeKinetics PDFaathiraAinda não há avaliações
- Michaelis Menten Equation SummaryDocumento7 páginasMichaelis Menten Equation SummaryBüşra GültekinAinda não há avaliações
- Enzyme KineticsDocumento8 páginasEnzyme KineticsronojoysenguptaAinda não há avaliações
- Enzyme M&LDocumento6 páginasEnzyme M&LNethra Sasikumar100% (1)
- FOOD MICROBIOLOGY Spoilage Poisoning and PreservationDocumento13 páginasFOOD MICROBIOLOGY Spoilage Poisoning and PreservationDr. SHIVA AITHAL100% (1)
- Hanuman Chalisa English MeaningDocumento3 páginasHanuman Chalisa English MeaningSaurabh SaxenaAinda não há avaliações
- 50 Books For ChildrenDocumento32 páginas50 Books For ChildrenDr. SHIVA AITHALAinda não há avaliações
- Microbiology of Water and Waste Water ManagementDocumento27 páginasMicrobiology of Water and Waste Water ManagementDr. SHIVA AITHAL100% (2)
- Air Microbiology 2009Documento13 páginasAir Microbiology 2009Dr. SHIVA AITHAL100% (9)
- Glycolysis and TCA Cycle Enzymes and EC Reference NumbersDocumento1 páginaGlycolysis and TCA Cycle Enzymes and EC Reference NumbersDr. SHIVA AITHALAinda não há avaliações
- Visit To ISDN BayalaluDocumento16 páginasVisit To ISDN BayalaluDr. SHIVA AITHAL100% (2)
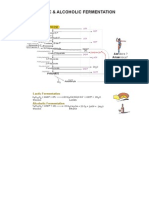
- Lactic and Alcoholic FermentationDocumento1 páginaLactic and Alcoholic FermentationDr. SHIVA AITHALAinda não há avaliações
- GlycolysisDocumento1 páginaGlycolysisDr. SHIVA AITHALAinda não há avaliações
- Structure of DNA by CrickDocumento8 páginasStructure of DNA by CrickDr. SHIVA AITHALAinda não há avaliações
- Modelo Doble Helice Watson y CrickDocumento2 páginasModelo Doble Helice Watson y Crickangelferp100% (2)
- Micro Teaching Skills ComponentsDocumento35 páginasMicro Teaching Skills ComponentsDr. SHIVA AITHAL89% (221)
- 3G Wireless NetworksDocumento16 páginas3G Wireless NetworksAli QureshiAinda não há avaliações
- A Brief Foray Into The World of Nano Materials by Sarbari BhattacharyaDocumento17 páginasA Brief Foray Into The World of Nano Materials by Sarbari BhattacharyaDr. SHIVA AITHALAinda não há avaliações
- Management of Libraries and Information Sciences by Naznin BanuDocumento12 páginasManagement of Libraries and Information Sciences by Naznin BanuDr. SHIVA AITHALAinda não há avaliações
- Preamble To The Constitution of India by Prema NagaleDocumento12 páginasPreamble To The Constitution of India by Prema NagaleDr. SHIVA AITHAL100% (1)
- Biotechnology in Agriculture Development by KS ManjunathDocumento15 páginasBiotechnology in Agriculture Development by KS ManjunathDr. SHIVA AITHAL100% (1)
- COA LAb File-ShubhiDocumento28 páginasCOA LAb File-ShubhiKrishan Kant ShuklaAinda não há avaliações
- The Effect of Axial Flow Velocity On Annular Gap Windage Power LossDocumento12 páginasThe Effect of Axial Flow Velocity On Annular Gap Windage Power Losskien nguyenAinda não há avaliações
- Stacks - Quesues and DequesDocumento23 páginasStacks - Quesues and DequesAbdallateef ShohdyAinda não há avaliações
- Seborg Chapter 1Documento3 páginasSeborg Chapter 1maykesguerraAinda não há avaliações
- Evaluating Capital ProjectsDocumento254 páginasEvaluating Capital Projectsariff_sodriAinda não há avaliações
- 3DC Real Light 24HDRi Vol03Documento27 páginas3DC Real Light 24HDRi Vol03AntezanaFernandoAinda não há avaliações
- Dell XPS 17 Quanta GM7 Rev D SchematicsDocumento39 páginasDell XPS 17 Quanta GM7 Rev D SchematicsvcompumatikAinda não há avaliações
- Science 3 PDFDocumento12 páginasScience 3 PDFJelyn CandoAinda não há avaliações
- MSC Academic Guide - Admission Test PDFDocumento13 páginasMSC Academic Guide - Admission Test PDFTonoy Peter CorrayaAinda não há avaliações
- 10) C2 Trigonometrical Identities and Equations QuestionsDocumento26 páginas10) C2 Trigonometrical Identities and Equations QuestionsBoxOneTwoAinda não há avaliações
- Measurement of Earthing Systems: Central Networks Earthing Manual Section E4Documento45 páginasMeasurement of Earthing Systems: Central Networks Earthing Manual Section E4ahmed_k7117Ainda não há avaliações
- Chapter 04 Chemical EquilibriumDocumento150 páginasChapter 04 Chemical Equilibriumnoel100% (6)
- CS 161 (Stanford, Winter 2024)Documento2 páginasCS 161 (Stanford, Winter 2024)Catriel LopezAinda não há avaliações
- AssignmentDocumento9 páginasAssignmentRazi BaigAinda não há avaliações
- Infinix NOTE 5 Stylus Quick Guide: Together We Can!Documento16 páginasInfinix NOTE 5 Stylus Quick Guide: Together We Can!LescribeurAinda não há avaliações
- Inserting Images - ShareLaTeX, Online LaTeX EditorDocumento11 páginasInserting Images - ShareLaTeX, Online LaTeX EditorAviraj3892Ainda não há avaliações
- IP03 Loops-4slides PDFDocumento10 páginasIP03 Loops-4slides PDFRamesh GAinda não há avaliações
- Sheet Pile Analysis Sheet v1.07-18.1Documento2 páginasSheet Pile Analysis Sheet v1.07-18.1SES DESIGN100% (1)
- Processing Sequence When Posting An EventDocumento5 páginasProcessing Sequence When Posting An EventSrinivas ManchiAinda não há avaliações
- Tutorial - How To Use The Poser Format ExporterDocumento7 páginasTutorial - How To Use The Poser Format ExporterMimhypopbummilha SwuAinda não há avaliações
- Caie As Computer Science 9618 Theory v3Documento20 páginasCaie As Computer Science 9618 Theory v3James HoangAinda não há avaliações
- Mechanics of MaterialsDocumento11 páginasMechanics of MaterialsMagesh KumarAinda não há avaliações
- Lec.5 (Prameters of OHTL Pt.2)Documento10 páginasLec.5 (Prameters of OHTL Pt.2)AbdullahAinda não há avaliações
- 111Documento10 páginas111Phước Ng50% (2)
- Mole Concept - L1rr PDFDocumento27 páginasMole Concept - L1rr PDFLegend KillerAinda não há avaliações
- IECEx INE 12.0010X 002Documento9 páginasIECEx INE 12.0010X 002Marcos SiqueiraAinda não há avaliações
- Question & Answers: Unity Solutions Specialist Exam For Implementation EngineersDocumento6 páginasQuestion & Answers: Unity Solutions Specialist Exam For Implementation EngineersAhmad Abd-dayeemAinda não há avaliações
- Pediatric Appendicitis ScoreDocumento6 páginasPediatric Appendicitis ScorePinandhito LatukolanAinda não há avaliações
- 5.4.1 EM Patch Release NotesDocumento11 páginas5.4.1 EM Patch Release Notessuraj saketAinda não há avaliações
- Clipping Clamping CircuitsDocumento7 páginasClipping Clamping CircuitsPavan SaiAinda não há avaliações