Você também pode gostar
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Random FactsDocumento353 páginasRandom FactsSergio Rivas100% (1)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- 6th Grade Spelling ListsDocumento10 páginas6th Grade Spelling Listsapi-326416336Ainda não há avaliações
- 56.vocal Warmup Log For Belt Your FaceDocumento5 páginas56.vocal Warmup Log For Belt Your FaceAlinutza AlinaAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- GEC PE 3 ModuleDocumento72 páginasGEC PE 3 ModuleMercy Anne EcatAinda não há avaliações
- MDB Specs PDFDocumento9 páginasMDB Specs PDFAbdElrahman AhmedAinda não há avaliações
- Functional Capacity Evaluation: Occupational Therapy's Role inDocumento2 páginasFunctional Capacity Evaluation: Occupational Therapy's Role inramesh babu100% (1)
- Fire Safety: Good Servant But A Bad MasterDocumento143 páginasFire Safety: Good Servant But A Bad MasterernmrajaAinda não há avaliações
- PLAB 2 VISA Guidelines by Omar AlamDocumento18 páginasPLAB 2 VISA Guidelines by Omar Alamrafew19Ainda não há avaliações
- Morita Therapy For Depression and AnxietyDocumento13 páginasMorita Therapy For Depression and AnxietyPedro GuimarãesAinda não há avaliações
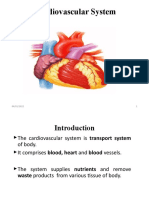
- Cardiovascular SystemDocumento47 páginasCardiovascular SystemAtnasiya GadisaAinda não há avaliações
- Reformulations and Branching Methods For Mixed-Integer Programming Chemical Production Scheduling ModelsDocumento10 páginasReformulations and Branching Methods For Mixed-Integer Programming Chemical Production Scheduling ModelsShamik MisraAinda não há avaliações
- On The Stability of Constrained MPC Without Terminal ConstraintDocumento5 páginasOn The Stability of Constrained MPC Without Terminal ConstraintShamik MisraAinda não há avaliações
- Search Engine Yearbook: Free Version:: March 2003Documento215 páginasSearch Engine Yearbook: Free Version:: March 2003Shamik MisraAinda não há avaliações
- D63 CBD 01Documento5 páginasD63 CBD 01Shamik MisraAinda não há avaliações
- ImmobilizationDocumento16 páginasImmobilizationDavid Muñoz HuachuhuillcaAinda não há avaliações
- F TestDocumento7 páginasF TestShamik MisraAinda não há avaliações
- Apport D Un Fonds de Commerce en SocieteDocumento28 páginasApport D Un Fonds de Commerce en SocieteJezebethAinda não há avaliações
- Plumber (General) - II R1 07jan2016 PDFDocumento18 páginasPlumber (General) - II R1 07jan2016 PDFykchandanAinda não há avaliações
- Neurovascular Assessment PDFDocumento5 páginasNeurovascular Assessment PDFNasrullah UllahAinda não há avaliações
- AIPMT 2009 ExamDocumento25 páginasAIPMT 2009 ExamMahesh ChavanAinda não há avaliações
- Cardiovascular Pharmacology: DR Muhamad Ali Sheikh Abdul Kader MD (Usm) MRCP (Uk) Cardiologist, Penang HospitalDocumento63 páginasCardiovascular Pharmacology: DR Muhamad Ali Sheikh Abdul Kader MD (Usm) MRCP (Uk) Cardiologist, Penang HospitalCvt RasulAinda não há avaliações
- Ens TecDocumento28 páginasEns TecBorja CanalsAinda não há avaliações
- Fermentation Media: Agustin Krisna WardaniDocumento27 páginasFermentation Media: Agustin Krisna WardaniYosiaAinda não há avaliações
- Thornburg Letter of RecDocumento1 páginaThornburg Letter of Recapi-511764033Ainda não há avaliações
- Evolution of Fluidized Bed TechnologyDocumento17 páginasEvolution of Fluidized Bed Technologyika yuliyani murtiharjonoAinda não há avaliações
- 2 Avaliação Edros 2023 - 7º Ano - ProvaDocumento32 páginas2 Avaliação Edros 2023 - 7º Ano - Provaleandro costaAinda não há avaliações
- Rekapan Belanja JKNDocumento5 páginasRekapan Belanja JKNAPOTEK PUSKESMAS MALEBERAinda não há avaliações
- Air Conditioning Is Inoperative, Fault Code 91eb11 or 91eb15 Is StoredDocumento5 páginasAir Conditioning Is Inoperative, Fault Code 91eb11 or 91eb15 Is Storedwasim AiniaAinda não há avaliações
- Muet Topic 10 City Life Suggested Answer and IdiomsDocumento3 páginasMuet Topic 10 City Life Suggested Answer and IdiomsMUHAMAD FAHMI BIN SHAMSUDDIN MoeAinda não há avaliações
- Reuse DNA Spin ColumnDocumento6 páginasReuse DNA Spin ColumnashueinAinda não há avaliações
- Coatings and Waterproofing: KhalafDocumento19 páginasCoatings and Waterproofing: KhalafmohamedAinda não há avaliações
- English Quarterly TestDocumento3 páginasEnglish Quarterly TestEdmon FabregasAinda não há avaliações
- Aphids and Ants, Mutualistic Species, Share A Mariner Element With An Unusual Location On Aphid Chromosomes - PMCDocumento2 páginasAphids and Ants, Mutualistic Species, Share A Mariner Element With An Unusual Location On Aphid Chromosomes - PMC2aliciast7Ainda não há avaliações
- Mapagbigay PT1 Group1Documento4 páginasMapagbigay PT1 Group1Hazel SarmientoAinda não há avaliações
- AST CatalogDocumento44 páginasAST CatalogCRISTIANAinda não há avaliações
- Child-Centered and Progressive EducationDocumento2 páginasChild-Centered and Progressive EducationDibyendu ChoudhuryAinda não há avaliações