Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Manganase Zinc FerritaDocumento11 páginasManganase Zinc FerritaFENFOGAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Application of Electrocoagulation Process For Dairy WastewaterDocumento9 páginasApplication of Electrocoagulation Process For Dairy WastewaterFENFOGAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Distinct 01X-MainDocumento9 páginasThe Distinct 01X-MainFENFOGAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (894)
- Emissions AthmosfericDocumento15 páginasEmissions AthmosfericFENFOGAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The INFLUENCE of NATURAL ORGANIC MATTER and AGING On SUSPENSION STABILITY in Guideline Toxicity Testing of Silver Zinc Oxide and Titanium Dioxide Nanopartcles With Daphania MagnaDocumento11 páginasThe INFLUENCE of NATURAL ORGANIC MATTER and AGING On SUSPENSION STABILITY in Guideline Toxicity Testing of Silver Zinc Oxide and Titanium Dioxide Nanopartcles With Daphania MagnaFENFOGAinda não há avaliações
- Investigation of MG (OH) AgentDocumento10 páginasInvestigation of MG (OH) AgentFENFOGAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Distinct 01X-MainDocumento9 páginasThe Distinct 01X-MainFENFOGAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Electrochemical Treatment and Operating Cost AnalysisDocumento11 páginasElectrochemical Treatment and Operating Cost AnalysisFENFOGAinda não há avaliações
- Eletro FloculaçãoDocumento6 páginasEletro FloculaçãoFENFOGAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The ContrainsDocumento7 páginasThe ContrainsFENFOGAinda não há avaliações
- Mechanistic Investigation Into Antibacterial Behaviour of Suspensions of ZnO Nanoparticles Against E CliDocumento13 páginasMechanistic Investigation Into Antibacterial Behaviour of Suspensions of ZnO Nanoparticles Against E CliFENFOGAinda não há avaliações
- Wate Manegenent 1 s2.0 S0956053X1400052X MainDocumento6 páginasWate Manegenent 1 s2.0 S0956053X1400052X MainFENFOGAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Green Synthetic Approach For Starch Capped Silver NanoparticlesDocumento10 páginasGreen Synthetic Approach For Starch Capped Silver NanoparticlesFENFOGAinda não há avaliações
- Silver Nanoparticles As Potential Antibacterial AgentsDocumento20 páginasSilver Nanoparticles As Potential Antibacterial AgentsFENFOGAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Review On Zinc ParticleDocumento15 páginasReview On Zinc ParticleFENFOGAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Biological Synthesis of Silver Nanoparticles and Evaluation of Antibacterial and Antifugal Properties of Silver and Copper NanoparticlesDocumento7 páginasBiological Synthesis of Silver Nanoparticles and Evaluation of Antibacterial and Antifugal Properties of Silver and Copper NanoparticlesFENFOGAinda não há avaliações
- Viscoelastic Analysis of Com-Plex Polymer Melt Flows Using The EXtended PomÔÇôPom Model - J.N.N.F.M 2002Documento26 páginasViscoelastic Analysis of Com-Plex Polymer Melt Flows Using The EXtended PomÔÇôPom Model - J.N.N.F.M 2002FENFOGAinda não há avaliações
- The Use of AntibacterialDocumento9 páginasThe Use of AntibacterialFENFOGAinda não há avaliações
- Modelling Film Formation and Degradation of Semi-Transparent Exterior Wood Coatings - 2007Documento12 páginasModelling Film Formation and Degradation of Semi-Transparent Exterior Wood Coatings - 2007FENFOGAinda não há avaliações
- Development of High Temperature-Resistant Hydrosoluble PaintDocumento1 páginaDevelopment of High Temperature-Resistant Hydrosoluble PaintFENFOGAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Reator CSTRDocumento5 páginasReator CSTRThiago SobreiraAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- Electrochemical Oxidation of A Textile Industry EffluentDocumento2 páginasElectrochemical Oxidation of A Textile Industry EffluentFENFOGAinda não há avaliações
- A Review of The Rheology of Filled Viscoelastic Systems - R.R 2003Documento36 páginasA Review of The Rheology of Filled Viscoelastic Systems - R.R 2003FENFOGAinda não há avaliações
- A New and Simple Procedure For The Evaluation of The Association of Surfactants To ProteinsDocumento8 páginasA New and Simple Procedure For The Evaluation of The Association of Surfactants To ProteinsFENFOGAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Laparoscopic MyomectomyDocumento2 páginasLaparoscopic MyomectomyclarissasveznaAinda não há avaliações
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDocumento7 páginasKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderAinda não há avaliações
- Methods, Method Verification and ValidationDocumento14 páginasMethods, Method Verification and Validationjljimenez1969100% (16)
- Optical Illusions and Vision Test AnswersDocumento8 páginasOptical Illusions and Vision Test AnswersZ'karia Al Ayi-zAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- FCPS SCERETS Rabia Ali Final - UPDATED Version 2014Documento9 páginasFCPS SCERETS Rabia Ali Final - UPDATED Version 2014Akas Rehman100% (4)
- Artificial Immunity Vs Natural ImmunityDocumento87 páginasArtificial Immunity Vs Natural ImmunityromeoqfacebookAinda não há avaliações
- Neural Network: Sudipta RoyDocumento25 páginasNeural Network: Sudipta RoymanAinda não há avaliações
- Effect of Gender On Short Term Memory FinishedDocumento27 páginasEffect of Gender On Short Term Memory Finishedhafsah286Ainda não há avaliações
- Respiration Course ObjectivesDocumento8 páginasRespiration Course Objectivesjoshy220996Ainda não há avaliações
- Cucumber Seed Germination: Effect and After-Effect of Temperature TreatmentsDocumento7 páginasCucumber Seed Germination: Effect and After-Effect of Temperature Treatmentsprotap uddin PolenAinda não há avaliações
- The Ecology of Photosintesis PathwaysDocumento5 páginasThe Ecology of Photosintesis PathwaysRicardo RicoAinda não há avaliações
- AP Bio Lab 1Documento5 páginasAP Bio Lab 1Kiran ShilaAinda não há avaliações
- Application of Edible Films and Coatings On MeatsDocumento2 páginasApplication of Edible Films and Coatings On MeatsAnasZeidAinda não há avaliações
- Exampro GCSE Biology: B2.5 InheritanceDocumento35 páginasExampro GCSE Biology: B2.5 InheritanceMeera ParaAinda não há avaliações
- Gen Bio 1 Module 1Documento30 páginasGen Bio 1 Module 1Louise Gabriel Lozada100% (1)
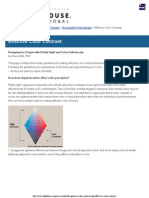
- Lighthouse International - Effective Color ContrastDocumento5 páginasLighthouse International - Effective Color ContrastVaishnavi JayakumarAinda não há avaliações
- Noise PollutionDocumento8 páginasNoise PollutiongoodthoughtsAinda não há avaliações
- Bio-Diversity Uses Threats and ConservationDocumento32 páginasBio-Diversity Uses Threats and ConservationRoshin VargheseAinda não há avaliações
- Specimen Cold Chain RequirementsDocumento6 páginasSpecimen Cold Chain RequirementsNikkae Angob0% (1)
- Chapter 1 of Blue MindDocumento23 páginasChapter 1 of Blue MindSouthern California Public Radio100% (1)
- Jhean Steffan Martines de CamargoDocumento2 páginasJhean Steffan Martines de CamargoJheanAinda não há avaliações
- Vol2 Final 19092016 PDFDocumento649 páginasVol2 Final 19092016 PDFshankarAinda não há avaliações
- Wbtet 2012Documento7 páginasWbtet 2012Partha BatabyalAinda não há avaliações
- Intergumentary System 10Documento40 páginasIntergumentary System 10Mike BasantaAinda não há avaliações
- Case Study On Dengue FeverDocumento76 páginasCase Study On Dengue FeverMary Rose Silva Gargar0% (1)
- Rankem SolventsDocumento6 páginasRankem SolventsAmmar MohsinAinda não há avaliações
- LAS 1 Cell TheoryDocumento12 páginasLAS 1 Cell TheoryJeremie CataggatanAinda não há avaliações
- Eosinophilic GastroenteritisDocumento7 páginasEosinophilic GastroenteritisSotir LakoAinda não há avaliações
- Post-Harvest Multiple Choice QuestionsDocumento8 páginasPost-Harvest Multiple Choice QuestionsErnestina OwireduAinda não há avaliações
- Plasma Gel Maker for Wrinkle Correction & Scar FillingDocumento1 páginaPlasma Gel Maker for Wrinkle Correction & Scar FillingPradeep AggarwalAinda não há avaliações
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeNo EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeNota: 4.5 de 5 estrelas4.5/5 (3)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeNo EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeNota: 5 de 5 estrelas5/5 (1)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolNo EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolAinda não há avaliações
- Napoleon's Buttons: 17 Molecules That Changed HistoryNo EverandNapoleon's Buttons: 17 Molecules That Changed HistoryNota: 4 de 5 estrelas4/5 (25)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeNo EverandChemistry for Breakfast: The Amazing Science of Everyday LifeNota: 4.5 de 5 estrelas4.5/5 (90)
- The Periodic Table: A Very Short IntroductionNo EverandThe Periodic Table: A Very Short IntroductionNota: 4.5 de 5 estrelas4.5/5 (3)