Você também pode gostar
- Scale, Heterogeneity, and the Structure and Diversity of Ecological CommunitiesNo EverandScale, Heterogeneity, and the Structure and Diversity of Ecological CommunitiesAinda não há avaliações
- Does God Roll Dice? Neutrality and Determinism in Evolutionary EcologyDocumento16 páginasDoes God Roll Dice? Neutrality and Determinism in Evolutionary EcologyAlara AğalarAinda não há avaliações
- RicklefsDocumento10 páginasRicklefsLeonardo UrruthAinda não há avaliações
- Ecological Niches and Geographic Distributions (MPB-49)No EverandEcological Niches and Geographic Distributions (MPB-49)Ainda não há avaliações
- Evolution in Changing Environments: Some Theoretical Explorations. (MPB-2)No EverandEvolution in Changing Environments: Some Theoretical Explorations. (MPB-2)Ainda não há avaliações
- Dynamics of Terrestrial GastropodsDocumento24 páginasDynamics of Terrestrial GastropodsDerrick Scott FullerAinda não há avaliações
- A Taxonomic Wish-List For Community Ecology: Nicholas J. GotelliDocumento13 páginasA Taxonomic Wish-List For Community Ecology: Nicholas J. GotelliMarlon Cezar CominettiAinda não há avaliações
- Statistical EcologyDocumento5 páginasStatistical EcologyjorgeAinda não há avaliações
- The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32)No EverandThe Unified Neutral Theory of Biodiversity and Biogeography (MPB-32)Nota: 5 de 5 estrelas5/5 (1)
- Field of Biogeography, Where Is Going - Avise, 2004Documento7 páginasField of Biogeography, Where Is Going - Avise, 2004DacanagaAinda não há avaliações
- Individual Size Variation Reduces Spatial Variation in Abundance of Tree Community Assemblage, Not of Tree PopulationsDocumento14 páginasIndividual Size Variation Reduces Spatial Variation in Abundance of Tree Community Assemblage, Not of Tree PopulationsRendi PermanaAinda não há avaliações
- A Unified Analysis of Niche Overlap Incorporating Data of Different TypesDocumento10 páginasA Unified Analysis of Niche Overlap Incorporating Data of Different TypesNelson AtencioAinda não há avaliações
- Ohlmann Et Al 2019 Div NetworksDocumento33 páginasOhlmann Et Al 2019 Div Networkskacagj bajazzoAinda não há avaliações
- Nabout Et Al 2010 - JZSERDocumento7 páginasNabout Et Al 2010 - JZSERjoaobioAinda não há avaliações
- N ICHE C ONSERVATISM: Integrating Evolution, Ecology, and Conservation BiologyDocumento23 páginasN ICHE C ONSERVATISM: Integrating Evolution, Ecology, and Conservation BiologyLaura Teresa PinzónAinda não há avaliações
- Landscape Ecology, Cross-Disciplinarity, and Sustainability ScienceDocumento4 páginasLandscape Ecology, Cross-Disciplinarity, and Sustainability ScienceThinh NguyenAinda não há avaliações
- Colwell Futuyma 1971 Ecology (Ancho de Nicho)Documento11 páginasColwell Futuyma 1971 Ecology (Ancho de Nicho)Carlos AlbertoAinda não há avaliações
- A Global Analysis of Cladoceran Body Size and Its Variation Linking To Habitat, Distribution and TaxonomyDocumento12 páginasA Global Analysis of Cladoceran Body Size and Its Variation Linking To Habitat, Distribution and TaxonomyBárbara BernardesAinda não há avaliações
- Ecological Society of AmericaDocumento14 páginasEcological Society of AmericaCleiton Lopes AguiarAinda não há avaliações
- Biological Diversity and Habitat Diversi PDFDocumento9 páginasBiological Diversity and Habitat Diversi PDFLuis Angel Aguirre MendezAinda não há avaliações
- FreshwB2006b PDFDocumento11 páginasFreshwB2006b PDFMiguel90Ainda não há avaliações
- Species ModelingDocumento20 páginasSpecies Modelingdominicrajkumar1Ainda não há avaliações
- Debinski and HoltDocumento14 páginasDebinski and HoltTaddese GashawAinda não há avaliações
- Sucesion y Colonizacion InglesDocumento10 páginasSucesion y Colonizacion InglesccbhAinda não há avaliações
- 5 Soberon 2009 Niches and Distributional Areas - Concepts, Methods, and AssumptionsDocumento7 páginas5 Soberon 2009 Niches and Distributional Areas - Concepts, Methods, and AssumptionsCesar ManriqueAinda não há avaliações
- Convergence and Divergence in A Long-Term Old-Field Succession: The Importance of Spatial Scale and Species AbundanceDocumento9 páginasConvergence and Divergence in A Long-Term Old-Field Succession: The Importance of Spatial Scale and Species AbundanceAlexis Centeno CuellarAinda não há avaliações
- Multivariate Legendre Etal EM2005Documento16 páginasMultivariate Legendre Etal EM2005Catalina Verdugo ArriagadaAinda não há avaliações
- Method and Theory of Catchment Analysis by Donna RopperDocumento23 páginasMethod and Theory of Catchment Analysis by Donna RopperKyle NormanAinda não há avaliações
- Addicott, Aho Et Al (OIKOS) 1987Documento7 páginasAddicott, Aho Et Al (OIKOS) 1987Dorina PllumbiAinda não há avaliações
- Niche Conservatism As An Emerging Principle in Ecology and Conservation BiologyDocumento15 páginasNiche Conservatism As An Emerging Principle in Ecology and Conservation BiologyagustinzarAinda não há avaliações
- Competition and the Structure of Bird Communities. (MPB-7), Volume 7No EverandCompetition and the Structure of Bird Communities. (MPB-7), Volume 7Nota: 3.5 de 5 estrelas3.5/5 (1)
- (Ricklefs, 2008) Disintegration of Community Concept PDFDocumento10 páginas(Ricklefs, 2008) Disintegration of Community Concept PDFOscar Leonardo Aaron Arizpe VicencioAinda não há avaliações
- 2007 Novack-Gottshall EcospaceDocumento32 páginas2007 Novack-Gottshall EcospaceobaytutAinda não há avaliações
- Hanski - 1998 - Metapopulation DynamicsDocumento10 páginasHanski - 1998 - Metapopulation DynamicszerainaitorAinda não há avaliações
- Soley-Guardia Et Al-2014-Journal of Biogeography PDFDocumento12 páginasSoley-Guardia Et Al-2014-Journal of Biogeography PDFSebas CirignoliAinda não há avaliações
- Relationships Between Population Densities and Niche-Centroid Distances in North American Birds - BDocumento10 páginasRelationships Between Population Densities and Niche-Centroid Distances in North American Birds - BnelsonatencioAinda não há avaliações
- Beyond Fragmentation - The Continuum Model For Fauna Research and Conservation inDocumento9 páginasBeyond Fragmentation - The Continuum Model For Fauna Research and Conservation inAna Carolina BritoAinda não há avaliações
- J Vegetation Science - 2013 - Correa Metrio - Environmental Determinism and Neutrality in Vegetation at Millennial Time PDFDocumento9 páginasJ Vegetation Science - 2013 - Correa Metrio - Environmental Determinism and Neutrality in Vegetation at Millennial Time PDFMariana Gutierrez MuneraAinda não há avaliações
- (Harrison & Cornell, 2008) Toward A Better Understanding of The Regional Causes of Local Community RichnessDocumento11 páginas(Harrison & Cornell, 2008) Toward A Better Understanding of The Regional Causes of Local Community RichnessOscar Leonardo Aaron Arizpe VicencioAinda não há avaliações
- Engen2008 PDFDocumento11 páginasEngen2008 PDFCamiBSchAinda não há avaliações
- Settlement Patterns and Ecosystem Pressures in the Peruvian Rainforest: Understanding the Impacts of Indigenous Peoples on BiodiversityNo EverandSettlement Patterns and Ecosystem Pressures in the Peruvian Rainforest: Understanding the Impacts of Indigenous Peoples on BiodiversityAinda não há avaliações
- Finding The Missing Link Between Landscape Structure and Population Dynamics: A Spatially Explicit PerspectiveDocumento23 páginasFinding The Missing Link Between Landscape Structure and Population Dynamics: A Spatially Explicit PerspectivemiramontesAinda não há avaliações
- DE LA VEGA, Gerardo J. & Pablo E. SchilmanDocumento8 páginasDE LA VEGA, Gerardo J. & Pablo E. SchilmanctiradoAinda não há avaliações
- L. A. S. Johnson Review No. 4 Bridging Historical and Ecological Approaches in BiogeographyDocumento10 páginasL. A. S. Johnson Review No. 4 Bridging Historical and Ecological Approaches in BiogeographyCecilia CorreaAinda não há avaliações
- Cadotte Etal GeneralitiesDocumento43 páginasCadotte Etal GeneralitiesMiguel90Ainda não há avaliações
- A Link Between Plant Traits and Abundance: Evidence From Coastal California Woody PlantsDocumento8 páginasA Link Between Plant Traits and Abundance: Evidence From Coastal California Woody PlantsLigia Paulillo SimsAinda não há avaliações
- Cavender Bares Etal2009Documento23 páginasCavender Bares Etal2009Loana AraujoAinda não há avaliações
- Historical Biogeography, Ecology and Species Richness: John J. Wiens and Michael J. DonoghueDocumento6 páginasHistorical Biogeography, Ecology and Species Richness: John J. Wiens and Michael J. DonoghuePatricia Matú ÁlvarezAinda não há avaliações
- Terrestrial Biomes A Conceptual ReviewDocumento13 páginasTerrestrial Biomes A Conceptual ReviewbrunoictiozooAinda não há avaliações
- Bio Divers I DadDocumento10 páginasBio Divers I Dadshaun6028Ainda não há avaliações
- Functional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewDocumento18 páginasFunctional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewrgelabertAinda não há avaliações
- Multilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceNo EverandMultilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceAinda não há avaliações
- Teoria de Nicho - ConceptosDocumento4 páginasTeoria de Nicho - ConceptosValeria AlvarezAinda não há avaliações
- Cadotte - Biol - Inv - 2006Documento13 páginasCadotte - Biol - Inv - 2006Miguel90Ainda não há avaliações
- 2020 - Lagomarsino & Frost - The Central Role of Taxonomy in The Study of Neotropical BiodeiversityDocumento17 páginas2020 - Lagomarsino & Frost - The Central Role of Taxonomy in The Study of Neotropical BiodeiversityArturoCosmasAvilaAinda não há avaliações
- From Populations to Ecosystems: Theoretical Foundations for a New Ecological Synthesis (MPB-46)No EverandFrom Populations to Ecosystems: Theoretical Foundations for a New Ecological Synthesis (MPB-46)Ainda não há avaliações
- Philo Q2 Lesson 5Documento4 páginasPhilo Q2 Lesson 5Julliana Patrice Angeles STEM 11 RUBYAinda não há avaliações
- Understanding The Contribution of HRM Bundles For Employee Outcomes Across The Life-SpanDocumento15 páginasUnderstanding The Contribution of HRM Bundles For Employee Outcomes Across The Life-SpanPhuong NgoAinda não há avaliações
- Cause Analysis For Spun Pile Crack and BrokenDocumento11 páginasCause Analysis For Spun Pile Crack and BrokenRony LesbtAinda não há avaliações
- L 11Documento3 páginasL 11trangAinda não há avaliações
- Mahindra&mahindraDocumento95 páginasMahindra&mahindraAshik R GowdaAinda não há avaliações
- G.Devendiran: Career ObjectiveDocumento2 páginasG.Devendiran: Career ObjectiveSadha SivamAinda não há avaliações
- Section 1 Company Overview StandardsDocumento34 páginasSection 1 Company Overview StandardsChris MedeirosAinda não há avaliações
- Radio Ac DecayDocumento34 páginasRadio Ac DecayQassem MohaidatAinda não há avaliações
- PDFDocumento653 páginasPDFconstantinAinda não há avaliações
- E7d61 139.new Directions in Race Ethnicity and CrimeDocumento208 páginasE7d61 139.new Directions in Race Ethnicity and CrimeFlia Rincon Garcia SoyGabyAinda não há avaliações
- Cap1 - Engineering in TimeDocumento12 páginasCap1 - Engineering in TimeHair Lopez100% (1)
- Manhole Head LossesDocumento11 páginasManhole Head Lossesjoseph_mscAinda não há avaliações
- Abilash - Subramanian CV - 003 PDFDocumento4 páginasAbilash - Subramanian CV - 003 PDFAbilash SubramanianAinda não há avaliações
- Government Hazi Muhammad Mohsin College Chattogram: Admission FormDocumento1 páginaGovernment Hazi Muhammad Mohsin College Chattogram: Admission FormThe Helper SoulAinda não há avaliações
- Engineering Ethics in Practice ShorterDocumento79 páginasEngineering Ethics in Practice ShorterPrashanta NaikAinda não há avaliações
- Ilovepdf MergedDocumento503 páginasIlovepdf MergedHemantAinda não há avaliações
- Agma MachineDocumento6 páginasAgma Machinemurali036Ainda não há avaliações
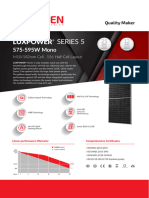
- Ficha Técnica Panel Solar 590W LuxenDocumento2 páginasFicha Técnica Panel Solar 590W LuxenyolmarcfAinda não há avaliações
- .CLP Delta - DVP-ES2 - EX2 - SS2 - SA2 - SX2 - SE&TP-Program - O - EN - 20130222 EDITADODocumento782 páginas.CLP Delta - DVP-ES2 - EX2 - SS2 - SA2 - SX2 - SE&TP-Program - O - EN - 20130222 EDITADOMarcelo JesusAinda não há avaliações
- Rankine-Froude Model: Blade Element Momentum Theory Is A Theory That Combines BothDocumento111 páginasRankine-Froude Model: Blade Element Momentum Theory Is A Theory That Combines BothphysicsAinda não há avaliações
- Thesis TipsDocumento57 páginasThesis TipsJohn Roldan BuhayAinda não há avaliações
- Project Name: Repair of Afam Vi Boiler (HRSG) Evaporator TubesDocumento12 páginasProject Name: Repair of Afam Vi Boiler (HRSG) Evaporator TubesLeann WeaverAinda não há avaliações
- Viscar Industrial Capacity LTD Company ProfileDocumento36 páginasViscar Industrial Capacity LTD Company ProfileTechnician MwangiAinda não há avaliações
- JO 20221109 NationalDocumento244 páginasJO 20221109 NationalMark Leo BejeminoAinda não há avaliações
- Press Statement - Book Launching Shariah Forensic 27 Oct 2023Documento4 páginasPress Statement - Book Launching Shariah Forensic 27 Oct 2023aAinda não há avaliações
- BIOAVAILABILITY AND BIOEQUIVALANCE STUDIES Final - PPTX'Documento32 páginasBIOAVAILABILITY AND BIOEQUIVALANCE STUDIES Final - PPTX'Md TayfuzzamanAinda não há avaliações
- Keeping Track of Your Time: Keep Track Challenge Welcome GuideDocumento1 páginaKeeping Track of Your Time: Keep Track Challenge Welcome GuideRizky NurdiansyahAinda não há avaliações
- Maximum and Minimum PDFDocumento3 páginasMaximum and Minimum PDFChai Usajai UsajaiAinda não há avaliações
- CHAPTER 2 Part2 csc159Documento26 páginasCHAPTER 2 Part2 csc159Wan Syazwan ImanAinda não há avaliações
- Dog & Kitten: XshaperDocumento17 páginasDog & Kitten: XshaperAll PrintAinda não há avaliações