Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Final NSCCBUE-226.4.2019-Seminar Abstract Book PDFDocumento120 páginasFinal NSCCBUE-226.4.2019-Seminar Abstract Book PDFManish KapoorAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Gel Electrophoresis PDFDocumento8 páginasGel Electrophoresis PDFManish KapoorAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Gel Electrophoresis PDFDocumento8 páginasGel Electrophoresis PDFManish KapoorAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Gel Electrophoresis PDFDocumento8 páginasGel Electrophoresis PDFManish KapoorAinda não há avaliações
- Climate 04 00008Documento23 páginasClimate 04 00008Manish KapoorAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Forage Seed TestingDocumento88 páginasForage Seed TestingManish KapoorAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Rabi 19-20Documento159 páginasRabi 19-20Manish KapoorAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Asparagus RapdDocumento8 páginasAsparagus RapdManish KapoorAinda não há avaliações
- Gulbenkian BGCI LRDocumento32 páginasGulbenkian BGCI LRManish KapoorAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- 84suINDUCED MUTAGENESIS IN CHRYSANTHEMUM MORIFOLIUM VARIETY OTOME PINK' THROUGH GAMMA IRRADIATIONp27 Kiran Kumari 2500 - FinalDocumento4 páginas84suINDUCED MUTAGENESIS IN CHRYSANTHEMUM MORIFOLIUM VARIETY OTOME PINK' THROUGH GAMMA IRRADIATIONp27 Kiran Kumari 2500 - FinalManish KapoorAinda não há avaliações
- LiliumDocumento9 páginasLiliumManish KapoorAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Book Abstracts Symposium 2008Documento207 páginasBook Abstracts Symposium 2008Manish KapoorAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Living Organisms Reproduce by Two Methods: Asexual ReproductionDocumento36 páginasLiving Organisms Reproduce by Two Methods: Asexual ReproductionMary SiddAinda não há avaliações
- BIOL 239 Lecture NotesDocumento39 páginasBIOL 239 Lecture NoteskcqywAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Gene StructureDocumento5 páginasGene StructureLia TuAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Name: - Date: - Period: - IB Biology - Meiosis Multiple Choice Practice Questions 1Documento5 páginasName: - Date: - Period: - IB Biology - Meiosis Multiple Choice Practice Questions 1Ghazal Fatima100% (2)
- Grade 8 Fourth Quarter First Semester NCR Living Things and Their EnvironmentDocumento6 páginasGrade 8 Fourth Quarter First Semester NCR Living Things and Their EnvironmentJohn SesconAinda não há avaliações
- Cell and Molecular Biology For Environmental EngineersDocumento44 páginasCell and Molecular Biology For Environmental EngineersCharleneKronstedtAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Biology Chapter 15Documento15 páginasBiology Chapter 15huifwiu jsbAinda não há avaliações
- ANAT2341 Embryology 2009 Lectures PDFDocumento232 páginasANAT2341 Embryology 2009 Lectures PDFTajamul islam100% (1)
- 6.3 MeiosisDocumento38 páginas6.3 Meiosiswickedbiology101Ainda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Meiosis ReportDocumento6 páginasMeiosis ReportDaizLee Ahmad71% (7)
- 5.4 Evolution - BioNinjaDocumento7 páginas5.4 Evolution - BioNinjaShuvi KurosakiAinda não há avaliações
- Concepts of Genetics 2nd Edition Brooker Solutions Manual 1Documento19 páginasConcepts of Genetics 2nd Edition Brooker Solutions Manual 1keithramseyfairdxtczw100% (24)
- Meiosis Worksheet 13 KeyDocumento2 páginasMeiosis Worksheet 13 KeyFarhan Pranadarmesta Merch100% (1)
- Karyotype Lab PDFDocumento5 páginasKaryotype Lab PDFwelfredo yuAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Cell Division Meiosis Homework AssignmentDocumento8 páginasCell Division Meiosis Homework Assignmenth00yfcc9100% (1)
- APEX Life ReproductionDocumento104 páginasAPEX Life Reproductionzerleigh dream pagalAinda não há avaliações
- These Are Diploid (2n) Ex. Blood Cells, Lung Cells, Muscle Cells, Heart Cells, Etc. These Are Haploid (N) Ex. Egg and SpermDocumento5 páginasThese Are Diploid (2n) Ex. Blood Cells, Lung Cells, Muscle Cells, Heart Cells, Etc. These Are Haploid (N) Ex. Egg and SpermLidia PrietoAinda não há avaliações
- Chapter 3 Cell DivisionDocumento5 páginasChapter 3 Cell DivisionNurul FasihahAinda não há avaliações
- Experiment No. 05: To Study Meiosis in Onion Bud CellsDocumento4 páginasExperiment No. 05: To Study Meiosis in Onion Bud CellsAryan Khode75% (4)
- HW 2Documento11 páginasHW 2John33% (30)
- Comparing Meiosis and MitosisDocumento6 páginasComparing Meiosis and MitosisGelo VeeAinda não há avaliações
- Cell Cycle and Cell Division - NotesDocumento9 páginasCell Cycle and Cell Division - NotesSaloni GuptaAinda não há avaliações
- Chapter 10 - MeiosisDocumento29 páginasChapter 10 - Meiosisnancie8Ainda não há avaliações
- Biology Chapter 6 Exercise (ANSWERS)Documento4 páginasBiology Chapter 6 Exercise (ANSWERS)NURUL AMIRA BINTI R AZMI KPM-GuruAinda não há avaliações
- Assignment-1: Class XIDocumento38 páginasAssignment-1: Class XIRinku VermaAinda não há avaliações
- BIOL 211 1 Roe Jflagg MC Practice Questions - SVDocumento5 páginasBIOL 211 1 Roe Jflagg MC Practice Questions - SVbmhshAinda não há avaliações
- LAS-Gen - Bio2 MELC 4 Q3-Week-2 PDFDocumento8 páginasLAS-Gen - Bio2 MELC 4 Q3-Week-2 PDFJayma RomeAinda não há avaliações
- Meiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inDocumento29 páginasMeiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inyamamaAinda não há avaliações
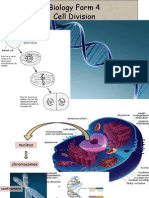
- Biology Form 4Documento13 páginasBiology Form 4Arvinth Guna SegaranAinda não há avaliações
- Cell DivisionDocumento5 páginasCell DivisionSsonko EdrineAinda não há avaliações