Você também pode gostar
- Método de Maxam y GilbertDocumento4 páginasMétodo de Maxam y GilbertElena ro alAinda não há avaliações
- Cuestionario Fisiologia Transporte de Oxigeno en La Sangre - RespiratorioDocumento2 páginasCuestionario Fisiologia Transporte de Oxigeno en La Sangre - RespiratorioIldaMercedesFriasGuerrero100% (1)
- Informe BioinformaticaDocumento15 páginasInforme Bioinformaticadiana medinaAinda não há avaliações
- Póster Glucólisis AnaeróbicaDocumento1 páginaPóster Glucólisis AnaeróbicaNaomi SacaAinda não há avaliações
- Locución para La Clase de EspermogramaDocumento7 páginasLocución para La Clase de EspermogramaElizabethAinda não há avaliações
- 10 Dna Mitocondrial y CloroplasticoDocumento5 páginas10 Dna Mitocondrial y Cloroplasticomandarina_sepsi9852100% (1)
- Clase Taller Tejido EpitelialDocumento4 páginasClase Taller Tejido EpitelialRogelio RiveroAinda não há avaliações
- Morfología de Los Eritrocitos2Documento6 páginasMorfología de Los Eritrocitos2Lorena ObandoAinda não há avaliações
- Capítulo 6 Nucleótidos y Ácidos NucleicosDocumento9 páginasCapítulo 6 Nucleótidos y Ácidos NucleicosAlfre PeAinda não há avaliações
- Taller de Replicacion y TraduccionDocumento4 páginasTaller de Replicacion y TraduccionNataliaCharryAinda não há avaliações
- Trabajo HemoglobinaDocumento3 páginasTrabajo HemoglobinaAlejandroToqueAinda não há avaliações
- Diseño de PrimersDocumento6 páginasDiseño de PrimersAdriana Herrera VivasAinda não há avaliações
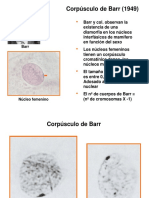
- Corpúsculo de BarrDocumento20 páginasCorpúsculo de BarrNoemi Chura0% (1)
- Modificaciones Postraduccionales y SecresiónDocumento15 páginasModificaciones Postraduccionales y SecresiónSeba CarnéAinda não há avaliações
- Cuestionario Embriologia IIDocumento5 páginasCuestionario Embriologia IILuis Alberto Solis RiveraAinda não há avaliações
- Técnica HistológicaDocumento11 páginasTécnica HistológicaAníbal CIAinda não há avaliações
- Preguntas Molecular Segundo CorteDocumento6 páginasPreguntas Molecular Segundo CorteValentina Arango100% (1)
- FOSFORODocumento1 páginaFOSFOROel zairAinda não há avaliações
- TRANSCRIPCIONDocumento20 páginasTRANSCRIPCIONJorgeLaTorreNavarroAinda não há avaliações
- Metabolismo de Nucleotidos.10Documento24 páginasMetabolismo de Nucleotidos.10Edwar Tarrillo Vasquez100% (1)
- Clonación Acelular PCRDocumento23 páginasClonación Acelular PCRVanessa Huirse LizanoAinda não há avaliações
- Extracción de ADN de Vegetales y FrutasDocumento2 páginasExtracción de ADN de Vegetales y FrutasDiegosr123Ainda não há avaliações
- Tincion de PASDocumento3 páginasTincion de PASRobert Ayala HerreraAinda não há avaliações
- EOSINÓFILOSDocumento5 páginasEOSINÓFILOSCarolina YeroviAinda não há avaliações
- Proteínas Estructura y FunciónDocumento37 páginasProteínas Estructura y FunciónPaola Duque100% (1)
- Informe Numero 5 Laboratorio (Discucion y Bibliografia)Documento4 páginasInforme Numero 5 Laboratorio (Discucion y Bibliografia)Carlos CarcamoAinda não há avaliações
- EsquemahematopoyesispdfDocumento3 páginasEsquemahematopoyesispdfmonchi026100% (1)
- Traduccion Feulgen KiernanDocumento4 páginasTraduccion Feulgen KiernanFrancisca Valentina Cárcamo UlloaAinda não há avaliações
- NUCLEÓTIDOSDocumento8 páginasNUCLEÓTIDOSCarlos Castilla EspinozaAinda não há avaliações
- Atlas de Histología (Tejido Epitelial)Documento18 páginasAtlas de Histología (Tejido Epitelial)José LópezAinda não há avaliações
- Adhesion CelularDocumento30 páginasAdhesion CelularBeto HdzAinda não há avaliações
- Bancos Bioquimica ExamenesDocumento21 páginasBancos Bioquimica Examenesmaricel gabriela acosta aguilarAinda não há avaliações
- Clase 11, Organelos CelularesDocumento25 páginasClase 11, Organelos CelularesCarolina Paz VillarroelAinda não há avaliações
- Retículo Endoplásmico Pi MZVDocumento29 páginasRetículo Endoplásmico Pi MZVCool BoyLVAinda não há avaliações
- Folleto Cobas 4800 PDFDocumento11 páginasFolleto Cobas 4800 PDFNoe Muñoz QuitoAinda não há avaliações
- Capitulo 27 StryerDocumento21 páginasCapitulo 27 StryerCristobal Ibarra SalazarAinda não há avaliações
- Bases NitrogenadasDocumento6 páginasBases NitrogenadasJohan MarquezAinda não há avaliações
- La CardiolipinaDocumento10 páginasLa Cardiolipinauniversal04-82100% (1)
- Cuantificación de ADN y ARN Por Espectrofotometría y Separación de ADN Por Electroforesis Horizontal en Gel de Agarosa.Documento21 páginasCuantificación de ADN y ARN Por Espectrofotometría y Separación de ADN Por Electroforesis Horizontal en Gel de Agarosa.Marien BlancoAinda não há avaliações
- Biologia Examen Junio 2012 CorregidoDocumento9 páginasBiologia Examen Junio 2012 CorregidoguillermoazorerohotmailcomAinda não há avaliações
- Extraccion Casera de AdnDocumento2 páginasExtraccion Casera de AdnKatia Aramburu SoteloAinda não há avaliações
- ReplicaciónDocumento6 páginasReplicaciónLucia HernandezAinda não há avaliações
- Introducción Extracción Del ADNDocumento2 páginasIntroducción Extracción Del ADNAlejandra TKTAinda não há avaliações
- Endosomas y LisosomasDocumento24 páginasEndosomas y LisosomasAurelia Gonzales Calle93% (14)
- CUESTIONES UT 7. Clonacion de AN 2020-21 RESUELTASDocumento3 páginasCUESTIONES UT 7. Clonacion de AN 2020-21 RESUELTASbanylexAinda não há avaliações
- Citogenética Convencional y Molecular - Nro de Bandas PDFDocumento38 páginasCitogenética Convencional y Molecular - Nro de Bandas PDFClaudia IpuchaAinda não há avaliações
- Prueba Adn 4Documento3 páginasPrueba Adn 4Antonella Tapia ArayaAinda não há avaliações
- Las Tecnicas y Tinciones EspecialesDocumento23 páginasLas Tecnicas y Tinciones EspecialesKaty100% (2)
- Preguntas 1er Parcial HistologíaDocumento8 páginasPreguntas 1er Parcial HistologíaTania RdzAinda não há avaliações
- Tinción para ReticulocitosDocumento6 páginasTinción para ReticulocitosJael TorrezAinda não há avaliações
- c6 Metabolismo de Las BacteriasDocumento30 páginasc6 Metabolismo de Las BacteriasMyle SaaAinda não há avaliações
- Test de TranscripcionDocumento3 páginasTest de TranscripcionAnonymous 7OTbV352GuAinda não há avaliações
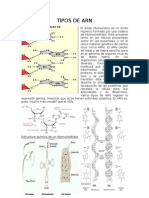
- Tipos de ArnDocumento5 páginasTipos de ArnMiguel Angel Rodas Herrera100% (2)
- HISTOLOGIADocumento41 páginasHISTOLOGIAPepe DelgadoAinda não há avaliações
- Cuerpos de Barr InformeDocumento9 páginasCuerpos de Barr Informejose coronado gonzalezAinda não há avaliações
- Estructura Del EritrocitoDocumento37 páginasEstructura Del EritrocitoMarco LojaAinda não há avaliações
- Sintesis de ProteinasDocumento7 páginasSintesis de ProteinasJaimecolina2004Ainda não há avaliações
- Eriptosis, La Muerte Suicida de Eritrocitos - Mecanismo y Enfermedades AsociadasDocumento11 páginasEriptosis, La Muerte Suicida de Eritrocitos - Mecanismo y Enfermedades Asociadasluthien5Ainda não há avaliações
- Replicación, Transcripción y TraducciónDocumento13 páginasReplicación, Transcripción y TraducciónPablo QuelalAinda não há avaliações
- Características de La Transcripción y Traducción GenéticaDocumento14 páginasCaracterísticas de La Transcripción y Traducción GenéticaSamuel Aldair0% (1)
- Resumen Final Biol. Cel. y Molec.Documento211 páginasResumen Final Biol. Cel. y Molec.Hugo MedusAinda não há avaliações
- REPLICACIONDocumento56 páginasREPLICACIONXimena CajamarcaAinda não há avaliações
- Junio 2021Documento9 páginasJunio 2021Sebas MamaAinda não há avaliações
- Unidad 1 y 2 - Mantenimiento y Variabilidad Del ADN NuclearDocumento7 páginasUnidad 1 y 2 - Mantenimiento y Variabilidad Del ADN NuclearEmma OchovaAinda não há avaliações
- Taller de ReplicaciónDocumento3 páginasTaller de ReplicaciónAmintaSusanaSarmientoUribeAinda não há avaliações
- BIOQUIMICADocumento93 páginasBIOQUIMICASanti VedoyaAinda não há avaliações
- GENETICADocumento22 páginasGENETICAKamy ChanAinda não há avaliações
- Control 6Documento2 páginasControl 6Tomás Ugarte EnrioneAinda não há avaliações
- Estructura Del Adn ProposicionesDocumento10 páginasEstructura Del Adn ProposicionesMaria Fernanda Rueda GuzmanAinda não há avaliações
- Ilovepdf MergedDocumento64 páginasIlovepdf MergedAndrea De La FuenteAinda não há avaliações
- Replicación - KarpDocumento14 páginasReplicación - KarpCuarto Paralelo 1100% (1)
- Clases Tema 10Documento12 páginasClases Tema 10UrielGranadosAinda não há avaliações
- Clase - 7 - Estructura y Composición Del DNA y ReplicaciónDocumento32 páginasClase - 7 - Estructura y Composición Del DNA y ReplicaciónJulian PoomAinda não há avaliações
- Organización Del Genoma y Flujo de Información en Bacterias (10-12)Documento15 páginasOrganización Del Genoma y Flujo de Información en Bacterias (10-12)ujbjhbAinda não há avaliações
- Tema 10 Replicación ADN MLDocumento38 páginasTema 10 Replicación ADN MLErnesto Lluesma DropoAinda não há avaliações
- Bloque 3 Biología: Genética y EvoluciónDocumento73 páginasBloque 3 Biología: Genética y EvoluciónJulia García-Ovies GutiérrezAinda não há avaliações
- Replicacion de ADNDocumento9 páginasReplicacion de ADNmamenmarinAinda não há avaliações
- Diferencias y Semejanzas Entre La Replicación Bacteriana y EucariotaDocumento2 páginasDiferencias y Semejanzas Entre La Replicación Bacteriana y EucariotaCristian EspinozaAinda não há avaliações
- Genomica EstructuralDocumento20 páginasGenomica EstructuralMística Epul SotoAinda não há avaliações
- Estructura Del ADN y Replicación Del ADNDocumento8 páginasEstructura Del ADN y Replicación Del ADNCamila LazzuriAinda não há avaliações
- Naturaleza Química de Los GenesDocumento11 páginasNaturaleza Química de Los GenesLitz Norma TFAinda não há avaliações
- Estructura Del ADN y ARN - GRADO 9Documento14 páginasEstructura Del ADN y ARN - GRADO 9Castro Abogados & AsesoresAinda não há avaliações
- Selectividad NucleotidosDocumento2 páginasSelectividad NucleotidosAmaia Teres LuriAinda não há avaliações
- Informe de Bioquimica DisertacionDocumento11 páginasInforme de Bioquimica DisertacionLuis GuzmánAinda não há avaliações
- 2008 Junio ADocumento2 páginas2008 Junio ASara SanabriaAinda não há avaliações
- Genetica Molecular EsquemaDocumento5 páginasGenetica Molecular Esquemaauladenaturales100% (2)
- Replicacion, Transcripcion y TraduccionDocumento3 páginasReplicacion, Transcripcion y TraduccionJorgelina PalacioAinda não há avaliações
- Tema 7. Genética Molecular.Documento11 páginasTema 7. Genética Molecular.Natalia Milla LopezAinda não há avaliações
- Transcripción Del ADN y TraducciónDocumento5 páginasTranscripción Del ADN y TraducciónSol HincapiéAinda não há avaliações