Você também pode gostar
- 911 Pigeon Disease & Treatment Protocols!No Everand911 Pigeon Disease & Treatment Protocols!Nota: 4 de 5 estrelas4/5 (1)
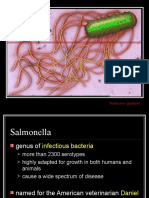
- Chapter-3 10 7-SalmonellosisDocumento20 páginasChapter-3 10 7-SalmonellosisSaulo EvangelistaAinda não há avaliações
- Salmon Ellos IsDocumento5 páginasSalmon Ellos IsIsak ShatikaAinda não há avaliações
- A Review On: Salmonellosis and Its Economic and Public Health SignificanceDocumento13 páginasA Review On: Salmonellosis and Its Economic and Public Health Significanceamanmalako50Ainda não há avaliações
- 07 For Shell 541554Documento14 páginas07 For Shell 541554frankyAinda não há avaliações
- II Bacterial DiseasesDocumento51 páginasII Bacterial DiseasesSagar AcharyaAinda não há avaliações
- Salmonellosis 1Documento15 páginasSalmonellosis 1BforbatAinda não há avaliações
- 1 See The Note in Chapter 2.9.8 Salmonellosis For The Principles Followed Concerning The Nomenclature of SalmonellaDocumento17 páginas1 See The Note in Chapter 2.9.8 Salmonellosis For The Principles Followed Concerning The Nomenclature of SalmonellaWira RamayadiAinda não há avaliações
- 2.03.11 Fowl TyphoidDocumento14 páginas2.03.11 Fowl TyphoidSaruediAinda não há avaliações
- Staphyloccocus Aureus PhatogenDocumento4 páginasStaphyloccocus Aureus PhatogenDaniel DelgadoAinda não há avaliações
- Chapter 9Documento12 páginasChapter 9Andi Dytha Pramitha SamAinda não há avaliações
- Rudolph PediatricDocumento9 páginasRudolph PediatricMuhammad RezaAinda não há avaliações
- 2.03.11 Fowl TyphoidDocumento11 páginas2.03.11 Fowl TyphoidYoni DarmawanAinda não há avaliações
- 375 FullDocumento11 páginas375 FullRenata YolandaAinda não há avaliações
- Genome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andDocumento7 páginasGenome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andArsh McBarsh MalhotraAinda não há avaliações
- SalmonellaDocumento8 páginasSalmonellaironAinda não há avaliações
- Table of ContentDocumento19 páginasTable of ContentasaAinda não há avaliações
- 146 - Salmonella SpeciesDocumento8 páginas146 - Salmonella SpeciesEmilio Emmanué Escobar CruzAinda não há avaliações
- International Journal of Food MicrobiologyDocumento7 páginasInternational Journal of Food MicrobiologyKatia RamónAinda não há avaliações
- 63 - 18 05 Isolation of Salm 220610Documento17 páginas63 - 18 05 Isolation of Salm 220610demasvebryAinda não há avaliações
- Case Study Salmonella in Caribbean FINALDocumento21 páginasCase Study Salmonella in Caribbean FINALGuia PurificacionAinda não há avaliações
- Antibiotic Resistance in Salmonella Spp. Isolated From Poultry: A GlobalDocumento15 páginasAntibiotic Resistance in Salmonella Spp. Isolated From Poultry: A GlobalKatia RamónAinda não há avaliações
- Bonardi., (2017)Documento14 páginasBonardi., (2017)ANGIE LORENA PALMA NI�OAinda não há avaliações
- Velgeetal 2012Documento17 páginasVelgeetal 2012shaznay delacruzAinda não há avaliações
- Paper 05Documento8 páginasPaper 05deltanueveAinda não há avaliações
- NIH Public Access: Author ManuscriptDocumento13 páginasNIH Public Access: Author ManuscriptJM TovarAinda não há avaliações
- Salmonella and ShigellaDocumento5 páginasSalmonella and ShigellaRigobertoAinda não há avaliações
- How Can We Solve The Problem of Salmonellosis in Poultry FarmDocumento13 páginasHow Can We Solve The Problem of Salmonellosis in Poultry FarmEvan HossainAinda não há avaliações
- FPUK-0714-61536364: Salmonella InfectionsDocumento11 páginasFPUK-0714-61536364: Salmonella InfectionsaliyahimranAinda não há avaliações
- BIO2107 - Tutorial 5 Discussion PointsDocumento4 páginasBIO2107 - Tutorial 5 Discussion PointsAjay Sookraj RamgolamAinda não há avaliações
- Salmonella: What Is Salmonella Infection (Salmonellosis) ?Documento9 páginasSalmonella: What Is Salmonella Infection (Salmonellosis) ?Bijay Kumar MahatoAinda não há avaliações
- Pathogenicity of SalmonellaDocumento22 páginasPathogenicity of SalmonellaTemidayoAinda não há avaliações
- Pediatrics in Review 2013 SalmonelosisDocumento11 páginasPediatrics in Review 2013 SalmonelosisCarlos Elio Polo VargasAinda não há avaliações
- Demam Tifoid 2016Documento8 páginasDemam Tifoid 2016Mona Mentari PagiAinda não há avaliações
- Tularemia: C H A P T E R 2 - 1 - 1 8Documento6 páginasTularemia: C H A P T E R 2 - 1 - 1 8WormInchAinda não há avaliações
- Typhoid FeverDocumento13 páginasTyphoid FeverPutri Nilam SariAinda não há avaliações
- 2.01.19 Vesicular StomitisDocumento11 páginas2.01.19 Vesicular StomitisWormInchAinda não há avaliações
- Food Safety HazardDocumento7 páginasFood Safety HazardPrithu GoelAinda não há avaliações
- "Presencia de Salmonella Spp. en Huevos Rosados para El ConsumoDocumento29 páginas"Presencia de Salmonella Spp. en Huevos Rosados para El ConsumoArturo Garcia GastañaduyAinda não há avaliações
- Tula Rae MiaDocumento38 páginasTula Rae Miashikha yadavAinda não há avaliações
- Salmonellosisincluding Entericfever: Farah Naz Qamar,, Wajid Hussain,, Sonia QureshiDocumento13 páginasSalmonellosisincluding Entericfever: Farah Naz Qamar,, Wajid Hussain,, Sonia QureshiAnak MuadzAinda não há avaliações
- Salmonella y BrucelosisDocumento19 páginasSalmonella y BrucelosisEdsonPerezAinda não há avaliações
- Incidence of Salmonella Spp. in Different Animal Species and Meat Products in Ecuador During The Period 2009-2019Documento8 páginasIncidence of Salmonella Spp. in Different Animal Species and Meat Products in Ecuador During The Period 2009-2019Deyaneira SanchezAinda não há avaliações
- Pui Et Al 2011Documento9 páginasPui Et Al 2011Muhammad Bilal Bin AmirAinda não há avaliações
- SalmonellaDocumento14 páginasSalmonellaTochukwu IbekweAinda não há avaliações
- Articulo 3 - SalmonellaDocumento16 páginasArticulo 3 - SalmonellaJoiver Alean FlórezAinda não há avaliações
- Salmonella, The Host and Disease: A Brief ReviewDocumento8 páginasSalmonella, The Host and Disease: A Brief ReviewPedro Albán MAinda não há avaliações
- SalmonellosisDocumento40 páginasSalmonellosispurposef49Ainda não há avaliações
- L15 SalmonellaDocumento19 páginasL15 SalmonellaMohammed RedhaAinda não há avaliações
- Unesco - Eolss Sample Chapters: Veterinary BacteriologyDocumento6 páginasUnesco - Eolss Sample Chapters: Veterinary BacteriologyGaluh EnggarAinda não há avaliações
- Flagellated Gram-Negative Bacterium Genus: SalmonellaDocumento1 páginaFlagellated Gram-Negative Bacterium Genus: SalmonellaMichael_William1990Ainda não há avaliações
- Uma Revisão Sobre Leptospitose BovinaDocumento9 páginasUma Revisão Sobre Leptospitose BovinaDiovana MachadoAinda não há avaliações
- Gram-Negative Rods Related To AnimalDocumento35 páginasGram-Negative Rods Related To AnimalAsa Mutia SAinda não há avaliações
- Salmonella 3Documento4 páginasSalmonella 3Jake MillerAinda não há avaliações
- Salmonella in Laying HenDocumento10 páginasSalmonella in Laying HenDaranee Nak-opatAinda não há avaliações
- Food InfectionDocumento20 páginasFood InfectiontesnaAinda não há avaliações
- Liver Fluke An Overview For PractitionersDocumento14 páginasLiver Fluke An Overview For PractitionersAli KareemAinda não há avaliações
- HGJK J LHB, JKDocumento4 páginasHGJK J LHB, JKChandra Noor RachmanAinda não há avaliações
- Animal Models of Salmonella Infections e PDFDocumento10 páginasAnimal Models of Salmonella Infections e PDFaminzaghiAinda não há avaliações
- (Lucian Boia) RomaniaDocumento331 páginas(Lucian Boia) Romanianela roxana100% (4)
- Mmymyy 113Documento9 páginasMmymyy 113frankyAinda não há avaliações
- Advances in Shrimp Aquaculture Management by Felix SDocumento95 páginasAdvances in Shrimp Aquaculture Management by Felix SfrankyAinda não há avaliações
- Guidelines For The Human Transportation of Research AnimalsDocumento2 páginasGuidelines For The Human Transportation of Research AnimalsfrankyAinda não há avaliações
- Transportation of Animals and Welfare: D.B. AdamsDocumento17 páginasTransportation of Animals and Welfare: D.B. AdamsfrankyAinda não há avaliações
- IATA-Live Animal RegulationsDocumento23 páginasIATA-Live Animal Regulationsfranky100% (1)
- Protocols For Transportation of Wild Animals: General Considerations Prior To Transport 1 Selection of IndividualsDocumento30 páginasProtocols For Transportation of Wild Animals: General Considerations Prior To Transport 1 Selection of IndividualsfrankyAinda não há avaliações
- Uterine and Vaginal Prolapse in EwesDocumento6 páginasUterine and Vaginal Prolapse in EwesfrankyAinda não há avaliações
- Complete Postparturient Uterine Prolapse in HF Cross Bred CowDocumento2 páginasComplete Postparturient Uterine Prolapse in HF Cross Bred CowfrankyAinda não há avaliações
- Welfare of Animals During Transport - Guidance NotesDocumento12 páginasWelfare of Animals During Transport - Guidance NotesfrankyAinda não há avaliações
- Fsa 3102Documento2 páginasFsa 3102frankyAinda não há avaliações
- Veterinarski Arhiv 82 (1), 11-24, 2012Documento14 páginasVeterinarski Arhiv 82 (1), 11-24, 2012frankyAinda não há avaliações
- Treatment Vaginal Prolapse PaperDocumento3 páginasTreatment Vaginal Prolapse PaperfrankyAinda não há avaliações
- Post Oestral Vaginal Prolapse in A Non-Pregnant Heifer (A Case Report)Documento7 páginasPost Oestral Vaginal Prolapse in A Non-Pregnant Heifer (A Case Report)frankyAinda não há avaliações
- I B Bu 201502004Documento7 páginasI B Bu 201502004frankyAinda não há avaliações
- Monitoring Postpartum Health in Dairy CowsDocumento7 páginasMonitoring Postpartum Health in Dairy CowsfrankyAinda não há avaliações
- Vet ArhivgenitalprolapseDocumento15 páginasVet ArhivgenitalprolapsefrankyAinda não há avaliações
- Uterine ProlapseDocumento3 páginasUterine ProlapsefrankyAinda não há avaliações
- 9 Unit 7 - Virology HODocumento90 páginas9 Unit 7 - Virology HOThirdy PinedaAinda não há avaliações
- Module 1 - EXPLOREDocumento3 páginasModule 1 - EXPLOREJoan BabieraAinda não há avaliações
- Biology ProjectDocumento18 páginasBiology ProjectParamdeep Singh100% (1)
- The Effects of COVID - 19 Pandemic Outbreak On Food Consumption Preferences and Their CausesDocumento6 páginasThe Effects of COVID - 19 Pandemic Outbreak On Food Consumption Preferences and Their CausesWahab Nurudeen OpeyemiAinda não há avaliações
- D-37/1, TTC MIDC, Turbhe, Navi Mumbai-400 703: ThyrocareDocumento3 páginasD-37/1, TTC MIDC, Turbhe, Navi Mumbai-400 703: ThyrocareSyed's Way PoolAinda não há avaliações
- Whistleblower (06.11.2023)Documento879 páginasWhistleblower (06.11.2023)tm8fq5xtbgAinda não há avaliações
- Administrative Order 2022-0010: Guidelines On TB-HIV Services Integration For Universal Health Care (UHC) ImplementationDocumento22 páginasAdministrative Order 2022-0010: Guidelines On TB-HIV Services Integration For Universal Health Care (UHC) ImplementationJovania B.Ainda não há avaliações
- Microbiology Unknown Lab ReportDocumento5 páginasMicrobiology Unknown Lab Reportfurban032780% (5)
- Reproduction in BacteriaDocumento20 páginasReproduction in BacteriaBharani Deepan100% (1)
- SHEA-APIC Guideline - Infection Prevention and Control in The Long-Term Care FacilityDocumento53 páginasSHEA-APIC Guideline - Infection Prevention and Control in The Long-Term Care FacilityAqsha RamadhanisaAinda não há avaliações
- Ficha Tecnica Divalash2Documento1 páginaFicha Tecnica Divalash2Paula CeledonAinda não há avaliações
- Expanded Program On Immunization ReportDocumento38 páginasExpanded Program On Immunization ReportKimm Delos ReyesAinda não há avaliações
- Toxin The - Cunning.of - Bacterial.poisons 0198605587Documento201 páginasToxin The - Cunning.of - Bacterial.poisons 0198605587Nicholas Woyshner100% (1)
- Submitted By: Deepali Talwar M.Tech (Environmental Engg.)Documento17 páginasSubmitted By: Deepali Talwar M.Tech (Environmental Engg.)deepalidtsAinda não há avaliações
- Primary SyphilisDocumento3 páginasPrimary SyphilisEqah TajuddinAinda não há avaliações
- Hesc 102Documento15 páginasHesc 102Aryansh AryanshAinda não há avaliações
- Stool AssessmentDocumento2 páginasStool AssessmentAlex MendozaAinda não há avaliações
- EMRA Antibiotic Guide EMRADocumento2 páginasEMRA Antibiotic Guide EMRAPaoNu0% (3)
- MICROPARADocumento61 páginasMICROPARAKyla RamonesAinda não há avaliações
- MCB 11L ReviewerDocumento1 páginaMCB 11L ReviewerJhoanna Mae MinionAinda não há avaliações
- Astm 2315Documento5 páginasAstm 2315yuanlupeAinda não há avaliações
- Control of Microorganisms in FoodsDocumento4 páginasControl of Microorganisms in FoodsMary KarimiAinda não há avaliações
- Fernando - Sico - Elo - (181111010) Bahasa InggrisDocumento2 páginasFernando - Sico - Elo - (181111010) Bahasa InggrisAsicoAinda não há avaliações
- 05-07-2020 HMB EnglishDocumento81 páginas05-07-2020 HMB Englishdhaneesh22Ainda não há avaliações
- ISK Pada Anak (Ade Sinta)Documento10 páginasISK Pada Anak (Ade Sinta)Ades SundayAinda não há avaliações
- Norfloxillin 200 Sol.: Bi D VW WJB 200 MJDocumento2 páginasNorfloxillin 200 Sol.: Bi D VW WJB 200 MJSharfina Akter BithiAinda não há avaliações
- How Should Colostrum-Deprived, Transport-Stressed Calves Be ManagedDocumento3 páginasHow Should Colostrum-Deprived, Transport-Stressed Calves Be Managedtaner_soysurenAinda não há avaliações
- Diagnosis and Management of Bacterial ConjunctivitisDocumento6 páginasDiagnosis and Management of Bacterial ConjunctivitisJonathan JoseAinda não há avaliações
- Microbiology 210: Final Laboratory ReportDocumento5 páginasMicrobiology 210: Final Laboratory ReportKcoope1788% (8)
- Parasitology: - IntroductionDocumento62 páginasParasitology: - IntroductionHana AliAinda não há avaliações
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessAinda não há avaliações
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNo EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNota: 4.5 de 5 estrelas4.5/5 (31)
- Will's Red Coat: The Story of One Old Dog Who Chose to Live AgainNo EverandWill's Red Coat: The Story of One Old Dog Who Chose to Live AgainNota: 4.5 de 5 estrelas4.5/5 (18)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNo EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueNota: 5 de 5 estrelas5/5 (4)
- An Eagle Named Freedom: My True Story of a Remarkable FriendshipNo EverandAn Eagle Named Freedom: My True Story of a Remarkable FriendshipAinda não há avaliações
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisNo EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisNota: 4 de 5 estrelas4/5 (9)
- The Other End of the Leash: Why We Do What We Do Around DogsNo EverandThe Other End of the Leash: Why We Do What We Do Around DogsNota: 5 de 5 estrelas5/5 (65)
- Uncontrolled Spread: Why COVID-19 Crushed Us and How We Can Defeat the Next PandemicNo EverandUncontrolled Spread: Why COVID-19 Crushed Us and How We Can Defeat the Next PandemicAinda não há avaliações
- The Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationNo EverandThe Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationNota: 4 de 5 estrelas4/5 (37)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsAinda não há avaliações
- Lucky Dog Lessons: Train Your Dog in 7 DaysNo EverandLucky Dog Lessons: Train Your Dog in 7 DaysNota: 4.5 de 5 estrelas4.5/5 (41)
- Do You Believe in Magic?: The Sense and Nonsense of Alternative MedicineNo EverandDo You Believe in Magic?: The Sense and Nonsense of Alternative MedicineAinda não há avaliações
- Show Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredNo EverandShow Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredNota: 3.5 de 5 estrelas3.5/5 (13)
- Come Back, Como: Winning the Heart of a Reluctant DogNo EverandCome Back, Como: Winning the Heart of a Reluctant DogNota: 3.5 de 5 estrelas3.5/5 (10)
- Your Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesNo EverandYour Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesNota: 4 de 5 estrelas4/5 (31)
- Second Chances: A Marine, His Dog, and Finding RedemptionNo EverandSecond Chances: A Marine, His Dog, and Finding RedemptionAinda não há avaliações
- Inside of a Dog: What Dogs See, Smell, and KnowNo EverandInside of a Dog: What Dogs See, Smell, and KnowNota: 4 de 5 estrelas4/5 (390)
- Edward's Menagerie: Dogs: 50 canine crochet patternsNo EverandEdward's Menagerie: Dogs: 50 canine crochet patternsNota: 3 de 5 estrelas3/5 (5)
- Dogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowNo EverandDogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowAinda não há avaliações
- The Wisdom of Plagues: Lessons from 25 Years of Covering PandemicsNo EverandThe Wisdom of Plagues: Lessons from 25 Years of Covering PandemicsNota: 4.5 de 5 estrelas4.5/5 (6)
- Our Dogs, Ourselves: The Story of a Singular BondNo EverandOur Dogs, Ourselves: The Story of a Singular BondNota: 4 de 5 estrelas4/5 (21)
- Animals Make Us Human: Creating the Best Life for AnimalsNo EverandAnimals Make Us Human: Creating the Best Life for AnimalsNota: 4.5 de 5 estrelas4.5/5 (2)
- I Am Bunny: How a ""Talking"" Dog Taught Me Everything I Need to Know About Being HumanNo EverandI Am Bunny: How a ""Talking"" Dog Taught Me Everything I Need to Know About Being HumanNota: 4.5 de 5 estrelas4.5/5 (8)