Você também pode gostar
- A 0 As 197 BDocumento12 páginasA 0 As 197 BqhqhqAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Endocrine System: University of SurreyDocumento13 páginasEndocrine System: University of SurreyqhqhqAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5795)
- Principles of Electrocardiography: Indiana University-Purdue UniversityDocumento11 páginasPrinciples of Electrocardiography: Indiana University-Purdue UniversityqhqhqAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- A Comparative Approach To Analysis and Modeling of Cardiovascular FunctionDocumento12 páginasA Comparative Approach To Analysis and Modeling of Cardiovascular FunctionqhqhqAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Physiological Modeling, Simulation, and Control: Joseph L. PalladinoDocumento30 páginasPhysiological Modeling, Simulation, and Control: Joseph L. PalladinoqhqhqAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- 01.watching ChildrenDocumento1 página01.watching ChildrenqhqhqAinda não há avaliações
- Q 2006 by Taylor & Francis Group, LLCDocumento14 páginasQ 2006 by Taylor & Francis Group, LLCqhqhqAinda não há avaliações
- 1 Data Management: Q 2006 by Taylor & Francis Group, LLCDocumento10 páginas1 Data Management: Q 2006 by Taylor & Francis Group, LLCqhqhqAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- The Biology and Biochemistry of Steroid Hormones: Advanced ArticleDocumento16 páginasThe Biology and Biochemistry of Steroid Hormones: Advanced ArticleqhqhqAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Wecb 652Documento8 páginasWecb 652qhqhqAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Systems Approach To Studying Disease: Advanced ArticleDocumento8 páginasSystems Approach To Studying Disease: Advanced ArticleqhqhqAinda não há avaliações
- Synthetic Chemistry: Formation of The Amide Bond: Advanced ArticleDocumento17 páginasSynthetic Chemistry: Formation of The Amide Bond: Advanced ArticleqhqhqAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Taste: Topics in Chemical Biology: Advanced ArticleDocumento12 páginasTaste: Topics in Chemical Biology: Advanced ArticleqhqhqAinda não há avaliações
- Auditory PathwayDocumento33 páginasAuditory PathwayAnusha Varanasi100% (1)
- UntitledDocumento378 páginasUntitledLaurent BarbierAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- 12b. The Central Auditory PathwayDocumento2 páginas12b. The Central Auditory PathwayShivan UmamaheswaranAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Physio HearingDocumento56 páginasPhysio Hearing65 Nidheesh Kumar PrabakamoorthyAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Acoustic Reflex Threshold (ART) Patterns: An Interpretation Guide For Students and Supervisors: Course Material and Exam Questions Course MaterialDocumento17 páginasAcoustic Reflex Threshold (ART) Patterns: An Interpretation Guide For Students and Supervisors: Course Material and Exam Questions Course Materialn_almagroAinda não há avaliações
- The Dangers of Edosing With Binaural Beats: PathwaysDocumento2 páginasThe Dangers of Edosing With Binaural Beats: PathwaysAgustina IturriAinda não há avaliações
- Health Aspects of Wireless Communication: Auditory Perception of Microwaves - Hearing Microwaves by James C. LinDocumento4 páginasHealth Aspects of Wireless Communication: Auditory Perception of Microwaves - Hearing Microwaves by James C. LinbingwazzupAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Foundations of Behavioral Neuroscience 9th Edition Carlson Test BankDocumento29 páginasFoundations of Behavioral Neuroscience 9th Edition Carlson Test Bankxeniaeira98897% (34)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Physics and Biology of AuditionDocumento100 páginasThe Physics and Biology of AuditionTAHA QAZIAinda não há avaliações
- Auditory PathwayDocumento16 páginasAuditory PathwayDrsreeram ValluriAinda não há avaliações
- James O. Pickles - An Introduction To The Physiology of Hearing-Emerald Group Publishing Limited (2012)Documento459 páginasJames O. Pickles - An Introduction To The Physiology of Hearing-Emerald Group Publishing Limited (2012)mane100% (1)
- 24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)Documento24 páginas24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)osamaeAinda não há avaliações
- NEURO Ch. 1-5 Quiz 1 NotesDocumento31 páginasNEURO Ch. 1-5 Quiz 1 NotesMelissa OrtizAinda não há avaliações
- Physiology of The Auditory SystemDocumento47 páginasPhysiology of The Auditory Systemmkct111Ainda não há avaliações
- Reflex Measurements: Quick GuideDocumento10 páginasReflex Measurements: Quick GuidefilippobAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Auditory PerceptionDocumento29 páginasAuditory PerceptionRose RodziAinda não há avaliações
- Masking Level Difference and ElectrophysiologicalDocumento8 páginasMasking Level Difference and ElectrophysiologicalFernanda CardozoAinda não há avaliações
- Hearing Physiology - GuytonDocumento11 páginasHearing Physiology - GuytonDuc HaAinda não há avaliações
- Translational Perspectives in Auditory Neuroscienc... - (7. The Superior Olivary Complex)Documento24 páginasTranslational Perspectives in Auditory Neuroscienc... - (7. The Superior Olivary Complex)Juliane NascimentoAinda não há avaliações
- Research PaperDocumento11 páginasResearch Paperapi-660335387Ainda não há avaliações
- Auditory ReceptorsDocumento15 páginasAuditory ReceptorsJeanette RiosAinda não há avaliações
- Acoustic - Stapedius - Reflexes PDFDocumento6 páginasAcoustic - Stapedius - Reflexes PDFSOFTMERCURYAinda não há avaliações
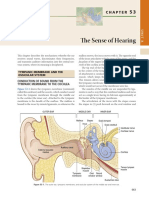
- The Sense of HearingDocumento11 páginasThe Sense of HearingAfdalia AmandaAinda não há avaliações
- Clin Aud I HandoutDocumento48 páginasClin Aud I HandoutelFadhlyAinda não há avaliações
- Auditory Pathways: Anatomy and Physiology: Handbook of Clinical Neurology March 2015Documento24 páginasAuditory Pathways: Anatomy and Physiology: Handbook of Clinical Neurology March 2015florensiaAinda não há avaliações
- The First Central Auditory Structure: The Cochlear NucleusDocumento56 páginasThe First Central Auditory Structure: The Cochlear NucleusAlison LuizAinda não há avaliações
- Sound Localization EssayDocumento11 páginasSound Localization EssayscholifyAinda não há avaliações
- Impedance Audiometry - Overview, Indications, ContraindicationsDocumento8 páginasImpedance Audiometry - Overview, Indications, ContraindicationsSorin AilincaiAinda não há avaliações
- Cap 11 - 22 Handbook of Clinical AudiologyDocumento216 páginasCap 11 - 22 Handbook of Clinical AudiologyCami IrribarraAinda não há avaliações
- 1 s2.0 S0306452214009932 MainDocumento12 páginas1 s2.0 S0306452214009932 MainRubén Alcalá ArgüellesAinda não há avaliações
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionNo EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionNota: 4 de 5 estrelas4/5 (404)