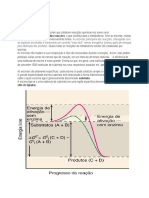
Você também pode gostar
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoAinda não há avaliações
- 2003 10 13168 Cinética e Propriedades EnzimáticasDocumento16 páginas2003 10 13168 Cinética e Propriedades EnzimáticasJoice CruzAinda não há avaliações
- Catalisadores BiológicosDocumento6 páginasCatalisadores BiológicosCleiton Belizario NetoAinda não há avaliações
- 1T.Cinética Química - 3ºanoEM.1 Etapa.2022Documento28 páginas1T.Cinética Química - 3ºanoEM.1 Etapa.2022Iara DaherAinda não há avaliações
- Estudo BioquimicaDocumento2 páginasEstudo Bioquimicadayannealves83% (6)
- EnzimasDocumento6 páginasEnzimasana_s_freireAinda não há avaliações
- Questionário enzimas: Fatores e tipos de inibiçãoDocumento9 páginasQuestionário enzimas: Fatores e tipos de inibiçãoRaffael Batista MarquesAinda não há avaliações
- Enzimas e processos fermentativosDocumento20 páginasEnzimas e processos fermentativosMatheus BañosAinda não há avaliações
- Enzimas bioengenhariaDocumento45 páginasEnzimas bioengenhariadaviAinda não há avaliações
- EnzimaDocumento11 páginasEnzimaMaestro VandaAinda não há avaliações
- Enzimas e reações metabólicasDocumento8 páginasEnzimas e reações metabólicasNareida MonteiroAinda não há avaliações
- JOSEFDocumento10 páginasJOSEFAnime BaAinda não há avaliações
- Enzimas e Metabolismo CelularDocumento10 páginasEnzimas e Metabolismo CelularNelson Pombal100% (2)
- EnzimasDocumento205 páginasEnzimasPâmela CristinaAinda não há avaliações
- EnzimasDocumento3 páginasEnzimaslucas022tgAinda não há avaliações
- Extração e Caracterização de EnzimasDocumento4 páginasExtração e Caracterização de EnzimasjoandroengAinda não há avaliações
- Trabalho de Biologia Celular2Documento9 páginasTrabalho de Biologia Celular2Jose PintoAinda não há avaliações
- Enzimas e Cinética EnzimáticaDocumento37 páginasEnzimas e Cinética EnzimáticamonekobeAinda não há avaliações
- Biologia 11 Classe I Trimestre 22 23Documento31 páginasBiologia 11 Classe I Trimestre 22 23jobhunhajosiasmatucaAinda não há avaliações
- ENEM 2019 Biologia Semana 07Documento190 páginasENEM 2019 Biologia Semana 07BlochMeisterAinda não há avaliações
- As enzimas: proteínas que catalisam reações bioquímicasDocumento5 páginasAs enzimas: proteínas que catalisam reações bioquímicasAngelique ViannyAinda não há avaliações
- EnzimasDocumento9 páginasEnzimasIsabellyAinda não há avaliações
- Bixo SP Semana 07Documento227 páginasBixo SP Semana 07kethelyn IsadoraAinda não há avaliações
- EnzimasDocumento15 páginasEnzimasines oliveiraAinda não há avaliações
- Fermentação e Atividade EnzimáticaDocumento36 páginasFermentação e Atividade EnzimáticaTatiana OliveiraAinda não há avaliações
- Colégio CAT: Enzimas e Metabolismo CelularDocumento30 páginasColégio CAT: Enzimas e Metabolismo CelularQuiala José100% (2)
- Resumo Expandido Atividade EnzimaticaDocumento4 páginasResumo Expandido Atividade EnzimaticaALLANAAinda não há avaliações
- Reação enzimática com pimentão verdeDocumento5 páginasReação enzimática com pimentão verdevitória souzaAinda não há avaliações
- Introdução À EnzimologiaDocumento23 páginasIntrodução À EnzimologiaFrancisco Oliveira100% (2)
- Estudo sobre enzimasDocumento4 páginasEstudo sobre enzimasVictor S. CarvalhoAinda não há avaliações
- En ZimasDocumento60 páginasEn ZimasEricaAinda não há avaliações
- 10 Eeq 5506Documento5 páginas10 Eeq 5506leandrohillenAinda não há avaliações
- Apostila EnzimologiaDocumento57 páginasApostila EnzimologiaBárbara MarinhoAinda não há avaliações
- Fermentação alcoólica e láctica: processos biológicos catalisados por enzimasDocumento1 páginaFermentação alcoólica e láctica: processos biológicos catalisados por enzimasInês RibeiroAinda não há avaliações
- Enzimas - SilvanDocumento39 páginasEnzimas - SilvanSilvandeOliveira50% (2)
- Composição Química da Célula - Enzimas como Catalisadores BiológicosDocumento3 páginasComposição Química da Célula - Enzimas como Catalisadores BiológicoslaviniaAinda não há avaliações
- 04 ProteinasDocumento5 páginas04 ProteinasSamuelSamuelAinda não há avaliações
- Atividade da enzima invertase livre determinada em diferentes temperaturasDocumento12 páginasAtividade da enzima invertase livre determinada em diferentes temperaturasPauloAinda não há avaliações
- Enzimas: propriedades e fatores que influenciam sua atividadeDocumento7 páginasEnzimas: propriedades e fatores que influenciam sua atividadeAline PiresAinda não há avaliações
- História e propriedades das enzimasDocumento12 páginasHistória e propriedades das enzimaskaratedoAinda não há avaliações
- EnzimasDocumento5 páginasEnzimasIsleia GomesAinda não há avaliações
- 06 EnzimasDocumento14 páginas06 EnzimasMatheus MAinda não há avaliações
- Estudo Dirigido 1 - BQA 2Documento7 páginasEstudo Dirigido 1 - BQA 2Esther LaisAinda não há avaliações
- EnzimasDocumento4 páginasEnzimasAlvim AlbertoAinda não há avaliações
- Aula EnzimasDocumento66 páginasAula EnzimasStefânya de Paula FreitasAinda não há avaliações
- Enzimas: estrutura, funções, regulação e classificaçãoDocumento3 páginasEnzimas: estrutura, funções, regulação e classificaçãoNayara SantiagoAinda não há avaliações
- Trabalho de Enzimas PDFDocumento5 páginasTrabalho de Enzimas PDFanon_387426005Ainda não há avaliações
- Curso Enem - Enzimas e proteínas especiaisDocumento180 páginasCurso Enem - Enzimas e proteínas especiaisDavi LacerdaAinda não há avaliações
- Relatorio Cinética Enzimática Da InvertaseDocumento20 páginasRelatorio Cinética Enzimática Da InvertaseJoaninha Faria100% (2)
- Atividade enzimática e fatores que influenciamDocumento11 páginasAtividade enzimática e fatores que influenciamLaís CaladoAinda não há avaliações
- Resumo BioquimicaDocumento22 páginasResumo BioquimicaedgardAinda não há avaliações
- Estudo Comparativo Da Atividade Da Amilase em Detergente em Pó e em SalivaDocumento14 páginasEstudo Comparativo Da Atividade Da Amilase em Detergente em Pó e em SalivaFelipe Pimentel PaixãoAinda não há avaliações
- Cinética Enzimática da Fosfatase AlcalinaDocumento4 páginasCinética Enzimática da Fosfatase AlcalinaLetícia CastroAinda não há avaliações
- Enzimas: Visão Geral e ClassificaçãoDocumento4 páginasEnzimas: Visão Geral e ClassificaçãoDébora Rocha100% (1)
- Estudo DirigidoDocumento5 páginasEstudo DirigidoJosé Rivaldo SoaresAinda não há avaliações
- Exercicio Parte Conceitual Do Capítulo de Enzimas - Janssen 10-03-2020Documento3 páginasExercicio Parte Conceitual Do Capítulo de Enzimas - Janssen 10-03-2020Janssen JulioAinda não há avaliações
- Enzimas: Uma breve introdução às enzimasDocumento19 páginasEnzimas: Uma breve introdução às enzimasjackspolAinda não há avaliações
- Enzimas catalíticasDocumento6 páginasEnzimas catalíticasAndré ScalaAinda não há avaliações
- Cinética Enzimática da Alfa-AmilaseDocumento15 páginasCinética Enzimática da Alfa-AmilaseJulie BarrosAinda não há avaliações
- Paleot 2 PDFDocumento14 páginasPaleot 2 PDFScribd_is_GreatAinda não há avaliações
- Dinossauros Andaram Por Aqui - Praia Da ParedeDocumento3 páginasDinossauros Andaram Por Aqui - Praia Da ParedeScribd_is_Great100% (1)
- PC Guia #246 PDFDocumento100 páginasPC Guia #246 PDFScribd_is_GreatAinda não há avaliações
- Departamento de Geologia da Universidade de LisboaDocumento19 páginasDepartamento de Geologia da Universidade de LisboaScribd_is_GreatAinda não há avaliações
- Equinodermes Do Cabeço Da Ladeira: Um Caso de Preservação Do Património GeológicoDocumento5 páginasEquinodermes Do Cabeço Da Ladeira: Um Caso de Preservação Do Património GeológicoScribd_is_GreatAinda não há avaliações
- Ficha Final Estudo Do MeioDocumento7 páginasFicha Final Estudo Do Meiodinamfaria100% (1)
- Grelhas Classificacao TestesDocumento6 páginasGrelhas Classificacao TestesXAinda não há avaliações
- Paleot 1Documento10 páginasPaleot 1Scribd_is_GreatAinda não há avaliações
- Ae 3ceb CN CoracaoDocumento1 páginaAe 3ceb CN CoracaoScribd_is_GreatAinda não há avaliações
- Aula5FRATURAS PDFDocumento63 páginasAula5FRATURAS PDFAlex RamiresAinda não há avaliações
- Grelha correção testes RFDocumento1 páginaGrelha correção testes RFScribd_is_GreatAinda não há avaliações
- Dinossáurios portugueses: pegadas reveladorasDocumento2 páginasDinossáurios portugueses: pegadas reveladorasScribd_is_GreatAinda não há avaliações
- Grelha correção testes RFDocumento1 páginaGrelha correção testes RFScribd_is_GreatAinda não há avaliações
- A Problemática Do Telemóvel Reflexão e ProcedimentosDocumento5 páginasA Problemática Do Telemóvel Reflexão e ProcedimentosScribd_is_GreatAinda não há avaliações
- 3º Ano-Ficha12Documento1 página3º Ano-Ficha12Carla Maria Pereira ViegasAinda não há avaliações
- 3º Ano AnimaisDocumento3 páginas3º Ano AnimaisScribd_is_GreatAinda não há avaliações
- Algas marinhas utilidadesDocumento19 páginasAlgas marinhas utilidadesYury LorenzAinda não há avaliações
- Omirante PTDocumento2 páginasOmirante PTScribd_is_GreatAinda não há avaliações
- Rochas e MineraisDocumento6 páginasRochas e MineraisAriana LimaAinda não há avaliações
- Minerais e Rochas MagmáticasDocumento2 páginasMinerais e Rochas MagmáticasRuiAlmeidaAinda não há avaliações
- Passeio Pedonal e Visita Guiada No Parque Natural Serra D'Aire e Candeeiros Dia 18 de MaioDocumento2 páginasPasseio Pedonal e Visita Guiada No Parque Natural Serra D'Aire e Candeeiros Dia 18 de MaioScribd_is_GreatAinda não há avaliações
- Texto de Visitação de Equinodermes para PNSACDocumento1 páginaTexto de Visitação de Equinodermes para PNSACScribd_is_GreatAinda não há avaliações
- João Varela Tese Mestrado Final PDFDocumento143 páginasJoão Varela Tese Mestrado Final PDFScribd_is_GreatAinda não há avaliações
- Ae 3oc Ciencias NaturaisDocumento13 páginasAe 3oc Ciencias NaturaisScribd_is_GreatAinda não há avaliações
- LVF Provas Escola ImgDocumento6 páginasLVF Provas Escola ImgScribd_is_GreatAinda não há avaliações
- 1-Formador Face Ao Contexto FuncoesDocumento70 páginas1-Formador Face Ao Contexto FuncoespmtoldaAinda não há avaliações
- Cor em Exames NacionaisDocumento22 páginasCor em Exames NacionaiscarlbispAinda não há avaliações
- Ler 1º Antes de InstalarDocumento1 páginaLer 1º Antes de InstalarScribd_is_GreatAinda não há avaliações
- Revistaecologia 6 Art 2 1Documento13 páginasRevistaecologia 6 Art 2 1Scribd_is_GreatAinda não há avaliações
- Dialnet ComoLocalizarOEpicentroDeUmTerremoto 5162227Documento13 páginasDialnet ComoLocalizarOEpicentroDeUmTerremoto 5162227Scribd_is_GreatAinda não há avaliações
- Manual Plataforma PantográficaDocumento39 páginasManual Plataforma Pantográficajesus.manhaesAinda não há avaliações
- Santillana - M11 - Ficha de Trabalho 5Documento3 páginasSantillana - M11 - Ficha de Trabalho 5Hugo SantosAinda não há avaliações
- Aulas 17 A 19 - Mquinas SncronasDocumento14 páginasAulas 17 A 19 - Mquinas SncronasAndré Soares da SilvaAinda não há avaliações
- Plano TCC - Rafael Figueiredo - r1Documento16 páginasPlano TCC - Rafael Figueiredo - r1Rafael FigueiredoAinda não há avaliações
- GlobmaticDocumento12 páginasGlobmaticPaulo Pinto100% (1)
- Cálculo da máxima altitude de foguetesDocumento4 páginasCálculo da máxima altitude de foguetesMARCUS ANDREY VIANAAinda não há avaliações
- Lagoas Anaeróbias: Tratamento de Esgotos com Baixo CustoDocumento60 páginasLagoas Anaeróbias: Tratamento de Esgotos com Baixo CustoFelipeAinda não há avaliações
- NR-12: Requisitos de segurança para máquinas e equipamentosDocumento88 páginasNR-12: Requisitos de segurança para máquinas e equipamentosFernando Martins100% (2)
- Captura de Tela 2023-02-04 À(s) 09.05.42Documento1 páginaCaptura de Tela 2023-02-04 À(s) 09.05.42laura diasAinda não há avaliações
- Vasco Moretto: Educar para Um Novo TempoDocumento7 páginasVasco Moretto: Educar para Um Novo TempoFelipe SantiagoAinda não há avaliações
- Centros Quadros e Paineis de DistribuicaoDocumento90 páginasCentros Quadros e Paineis de DistribuicaoCaldeira ElétricaAinda não há avaliações
- Derivadas de funções compostas e implícitasDocumento5 páginasDerivadas de funções compostas e implícitasPedro HenriqueAinda não há avaliações
- Catalogo-Supervisao-REV-14.12.23Documento22 páginasCatalogo-Supervisao-REV-14.12.23ASTEC PERAinda não há avaliações
- FrasesDocumento4 páginasFrasesAlex PiratiniAinda não há avaliações
- Algebra - 2 Fase Dos Vestibulares 2 (2018)Documento6 páginasAlgebra - 2 Fase Dos Vestibulares 2 (2018)Luiz Antonio Ponce AlonsoAinda não há avaliações
- Apostila Concreto ArmadoDocumento159 páginasApostila Concreto ArmadoLuciano RibeiroAinda não há avaliações
- 443Documento8 páginas443Júlio CésarAinda não há avaliações
- Apostila Refrigeração IndustrialDocumento41 páginasApostila Refrigeração IndustrialNelson Silva100% (2)
- Ficha Tecnica Icf Plastbau ParedesDocumento2 páginasFicha Tecnica Icf Plastbau ParedesPedroJorge_UCAinda não há avaliações
- Introdução A Química Exercícios - PDF 0Documento55 páginasIntrodução A Química Exercícios - PDF 0Fabiano AlvesAinda não há avaliações
- Moyses v2c08Documento11 páginasMoyses v2c08EphramoliverAinda não há avaliações
- Relatório Experimento 4Documento5 páginasRelatório Experimento 4Eduarda LealAinda não há avaliações
- Lista de exercícios de dinâmicaDocumento2 páginasLista de exercícios de dinâmicaHenry Moreno0% (1)
- Punto BRDocumento235 páginasPunto BRduhkacttusAinda não há avaliações
- Análise de circuitos elétricosDocumento6 páginasAnálise de circuitos elétricosmscaboraAinda não há avaliações
- Exercícios Experimento1Documento3 páginasExercícios Experimento1thayna.borgesAinda não há avaliações
- Aula 5 - Exercícios Tabela Periódica e Propriedades PeriódicasDocumento3 páginasAula 5 - Exercícios Tabela Periódica e Propriedades PeriódicasnatyrrrAinda não há avaliações
- Semicondutores: materiais e dispositivos básicosDocumento32 páginasSemicondutores: materiais e dispositivos básicosrewtwAinda não há avaliações
- Física 2 - Calor e Sua PropagaçãoDocumento2 páginasFísica 2 - Calor e Sua PropagaçãoArret Lorak100% (1)
- QI II Compostos de Coordenação III-TCCDocumento46 páginasQI II Compostos de Coordenação III-TCCLuanAinda não há avaliações