[Cell Cycle 7:3, 1-7; 1 February 2008]; 2008 Landes Bioscience
The calcineurin/NFAT signaling pathway is unique to verte-
brates and clear genetic evidences show that it plays critical
roles in orchestrating the intricate cellular interactions that
characterize vertebrate development and morphogenesis. In this
setting, the transcriptional regulators of the NFAT family func-
tion as molecular integrators of specific calcium signals with other
signaling pathways, including MAPkinase, WNT or NOTCH.
Deregulation of calcineurin/NFAT signaling and/or abnormal
expression of its components have recently been reported in solid
tumors of epithelial origin, lymphoma and lymphoid leukemia.
Our studies in mouse models of human T-ALL/lymphoma
shows that persistent activation of calcineurin/NFAT signaling
is pro-oncogenic in vivo and can be efficiently targeted by well-
characterized calcineurin inhibitors. We further discuss facts
and hypotheses concerning the molecular events that may act
upstream and downstream of calcineurin and/or NFAT activation
in different type of cancer cells.
The Calcineurin/NFAT Signaling Pathway
Calcineurin (PP2B) is a calcium-calmodulin-dependent serine/
threonine phosphatase implicated in a number of biological processes
(reviewed in refs. 13). In vertebrates, calcineurin is a heterodimer
composed of a catalytic subunit A (CnA, 5962kDa) and a regula-
tory subunit B (CnB, 19kDa). Three genes encoding the catalytic
subunits have been described in vertebrates. CnA and are
expressed ubiquitously whereas CnA expression is restricted to testis
and brain.
2,4,5
In addition to the phosphatase catalytic domain, CnA
contains a CnB-binding domain, a calmodulin-binding domain
as well as a carboxy-terminal autoinhibitory domain (Fig. 1). The
calcineurin regulatory subunit is encoded by two genes: CnB2,
which is specifically expressed in testis and CnB1, which exhibits an
ubiquitous expression pattern. In mice, deletion of the CnB1 gene
completely impairs calcineurin enzymatic activity in somatic tissues
and results in embryonic lethality at day 11 of development due to
severe defects in vascular patterning.
6
Under physiological conditions, engagement of cell-surface recep-
tors coupled to phospholipase C activation (e.g., the antigen receptor
in mature T and B cells) results in the generation of Inositol-
(1,4,5)trisphosphate (InsP3) and diacylglycerol (DAG). While DAG
activates the RAS/PKC pathway, InsP3 mediates the release of calcium
from internal stores, which in turn induces the opening of specific
store-operated calcium channels (CRAC). This results in the influx
of extracellular calcium and the calcium/calmodulin-dependent
activation of calcineurin (Fig. 1; reviewed in ref. 7) and the ensuing
dephosphorylation of its substrates, including NFAT (Nuclear Factor
of Activated T cell) proteins (Fig. 2). NFAT is a family of 5 members:
NFATc1, c2, c3, c4 and NFAT5, the latter being the only family
member not regulated by calcineurin. Mouse genetic studies have
demonstrated a strong epistatic relationship between calcineurin and
NFATs activation and function in many developmental processes
(reviewed in refs. 6 and 8 and references therein).
Perspective
The calcineurin/NFAT signaling pathway
A novel therapeutic target in leukemia and solid tumors
Hind Medyouf and Jacques Ghysdael*
CNRS UMR146 and Institut Curie; Centre Universitaire; Orsay, France
Key words: calcineurin, NFAT, leukemia, lymphoma, solid tumors
*Correspondence to: Jacques Ghysdael; CNRS UMR146 and Institut Curie;
Centre Universitaire; Bat 110; Orsay 91405 France; Tel.: 33.1.69863152; Fax:
33.1.69.07.45.25; Email: Jacques.Ghysdael@curie.u-psud.fr
Submitted: 11/05/07; Revised: 11/21/07; Accepted: 11/23/07
Previously published online as a Cell Cycle E-publication:
http://www.landesbioscience.com/journals/cc/article/5357
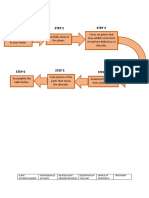
Figure 1. Mechanism of calcineurin activation. Caleincurin is composed of a
catalytic (CnA) and a regulatory (CnB) subunit. In unstimulated cells, calmod-
ulin is not associated with calcincurin and the CnA C-terminal autoinhibi-
tory domain (AID) interacts with the catalytics cleft and inhibits calcineurin
phosphatase activity. Signal-evoked increase in intracellular calcuim results
in calcium-dependent binding of calmodulin to calcineurin, thus relieving the
inhibitory activity of the AID on the catalytics domain and resulting in the
dephosphorylation of calcineurin substrates, including NFAT.
www.landesbioscience.com Cell Cycle 297
The calcineurin/NFAT signaling pathway in cancer
NFATc1, c2 and c3 are the only members expressed in the
lymphoid lineage. In resting lymphocytes, NFAT is located in the
cytoplasm as a hyperphosphorylated, inactive form. Under these
conditions, NFAT phosphorylation is insured by the combined
action of several maintenance kinases, including CK1 and DYRK2
that target specific serine residues in the NFAT conserved regulatory
domain. Signaling through calcium/calcineurin results in NFAT
proteins dephosphorylation, causing a conformational switch that
unmasks their nuclear localization sequence (NLS) and allows their
translocation to the nucleus, where they bind to specific DNA
response elements to regulate transcription in synergy with a number
of other transcriptional regulators. Mice with conditional deletion
of the CnB1 gene early in thymocytes development exhibit an 80%
decrease in thymic cellularity, perturbed pre-TCR signaling and the
complete absence of TCR-mediated positive selection of mature T
cells.
9
The importance of calcineurin downstream of TCR signaling
in peripheral T cells is underscored by the fact that (1) two effective
immunosuppressant used in human medicine,
FK506 and cyclosporine A (CsA) are inhibitors
of calcineurin; (2) patients with a rare form of
hereditary severe combined immunodeficiency
(SCID) show a selective defect in calcineurin/
NFAT activation.
10
Targeted deletion of CnB1
in B cells shows an essential role for calcineurin
in cell proliferation in response to in vitro anti-
IgM stimulation of the antigen receptor (BCR).
11
In contrast, B cell responses in vivo are partially
independent of calcineurin, likely due to the inte-
gration of BCR signaling with other cell surface
receptor signals. This study also showed that self
antigen-mediated selection of the B1 cell subset is
entirely dependent upon calcineurin.
11
Calcineurin and NFAT Factors in Cancer
Calcineurin and/or its downstream NFAT
targets have recently been implicated in cancer.
12
Tissue sections from invasive ductal breast carci-
noma patients display high expression of NFATc2
and NFAT5 in tumor cells.
13
Studies in breast
carcinoma cell lines show that NFAT expres-
sion and transcriptional activation are induced
downstream of integrin signaling, are indepen-
dent of calcineurin activity and promote cell
migration and matrigel invasion, suggesting a
role for NFATc2/NFAT5 in carcinoma invasive-
ness in vivo.
13
Buchholz and colleagues have
shown that about 70% of pancreatic carcinoma
show high level expression of nuclear NFATc1 as
compared to healthy pancreatic tissue.
14
Using
human-derived pancreatic carcinomas cell lines,
the authors demonstrated that the nuclear local-
ization of transcriptionally active NFATc1 is a
calcineurin-dependent process since it was inhib-
ited by CsA. Treatment with CsA also inhibited
in vitro cell cycle progression and anchorage-inde-
pendent proliferation of the Panc1 cell line.
In addition to their proposed implication in solid
tumors, the involvement of calcineurin and NFAT is also suspected
in hematologic malignancies. Marafioti et al. surveyed a large panel of
Non Hodgkin B-cell and T-cell lymphomas for NFATc1 expression
and nuclear localization.
15
Nuclear localization of NFATc1 was found
in 70% of the cases of Burkitt lymphoma (BL) and about 30% of the
cases of diffuse large B cell lymphoma (DLBCL). Nuclear localiza-
tion of NFATc1 or dephosphorylation of both NFATc1 and NFATc2
was also observed in DLBCL patient material
16,17
and in aggressive
T cell lymphoma.
17
NFAT activation in cell lines derived from
DLBCL and T cell acute lymphoblastic leukemia (T-ALL) is calci-
neurin-dependent as it was suppressed in response to CsA or FK506
treatment.
16,17
CsA treatment inhibited cell cycle progression and
induced apoptosis in these lines. The in vivo relevance of these
observations was obtained in mouse models of human T-ALL/
lymphoma induced either by constitutive activation of the JAK/
STAT or NOTCH1 signaling pathways. In both systems, sustained
activation of the calcineurin/NFAT signaling module was observed
Figure 2. Schematic view of the calcineurin/NFAT signaling pathway. Engagement by their
ligand of cell surface receptors coupled to the activation of phospholipase C or phospholipase
C results in the hydrolysis of phophatidylinositol 4,5 bisphosphate (PIP2) into diacylglycerol
(DAG) and inositol 1,4,5 triphosphate (InsP3). InsP3 binds to a receptor in the endoplasmic
reticulum (ER) to release calcium ions stored in the ER. Store depletion results in the opening of
calcium release-activated channels at the plasma membrane and in the import of extracellular
calcium. The resulting increase in intracellular calcium activates calmodulin and a series of
calmodulin-dependent enzymes, including the protein phosphatase calcineurin (Cn). Caclium- and
calmodulin-activated calcineurin dephosphorylates the 1214 serine residue in NFAT regulatory
domain that are constitutively phosphorylated in quiescent cells. Phosphorylated NFAT is cyto-
solic and its calcium-mediated dephosphorylation by calcineurin leads to a concerted change in
conformation, leading to its nuclear translocation. Nuclear NFAT cooperates with a number of
other transcriptional regulators to integrate calcium signaling events with other signaling inputs at
the transcriptional level. Cycolsporine A (CsA) and FK506 (Tacrolimus) are unrelated compounds
that inhibit calcineurin activity towards its protein substrates following their interactions with spe-
cific immunophilins, namely cyclophilin A and FKBP12.
298 Cell Cycle 2008; Vol. 7 Issue 3
The calcineurin/NFAT signaling pathway in cancer
in leukemic cells and treatment of diseased mice with either CsA or
FK506 resulted in leukemia regression linked to inhibition of tumor
cell proliferation, induction of apoptosis and stoechiometric NFAT
rephosphorylation.
17
Moreover, transduction of leukemic cells with a
calcium-independent, constitutively activated mutant of calcineurin
was found to enhance their aggressiveness and to enhance leukemia
progression in vivo, consistent with an intrinsic requirement for
calcineurin in leukemic cells. It is possible that treatment of leukemic
mice with CsA or FK506 also inhibited a function(s) in cells of the
tumor microenvironment (e.g., bone marrow stromal cells; blood
vessel cells) that could assist leukemic cell proliferation or survival.
Taken together, these data provide compelling evidence that calci-
neurin activation contributes to the pathogenesis of T-ALL and other
aggressive lymphoid malignancies.
NFAT Proteins as Potential Mediators of Calcineurin Activity
in Cancer
Members of the NFAT family are prominent targets of calcineurin
which are involved in the regulation of a large number of genes crit-
ical for proliferation, growth, migration, differentiation and survival
of cells in many lineages (reviewed in ref. 3). Since deregulation of
these phenotypic traits is commonly observed in cancer cells, an
oncogenic potential for the NFAT proteins has long been suspected
but only recently reported.
Calcium/calcineurin/NFAT signaling was discovered as an essen-
tial pathway acting downstream of the TCR to induce expression
of cytokine genes and drive proliferation of initially quiescent T
cells.
18,19
Furthermore, the importance of this signaling pathway to
cell cycle progression, in particular at the G
1
-to-S transition is well
established (reviewed in refs. 20 and 21). In line with this, proteins
of the NFAT family have been shown to directly regulate positively
or negatively the expression of genes implicated in cell cycle control,
including CDK4,
22
cyclin A2
23
and p21 (WAF/Cip1).
24
Moreover
NFAT loss-of-function or gain-of-function leads to deregulation of
D-type and E-type cyclins expression in several cell types, including
lymphocytes.
11,25,26
The calcineurin/NFAT signaling module is
also involved in cell survival as exemplified by the increased apop-
tosis observed in developing DP thymocytes of NFATc3-deficient
mice,
27,28
but the molecular mechanisms involved are still debated.
Importantly, although acting downstream from well characterized
pro-survival factors such as neurotrophins in neuronal cells, the
calcineurin/NFAT pathway is not involved in survival of these cells,
indicating clear differences depending upon the cellular context.
29
During normal development, the calcineurin/NFAT signaling
module plays a critical role in vasculogenesis and angiogenesis and in
the regulation of the VEGF pathway
30-32
and its abnormal deregula-
tion in cancer cells could be involved in tumor neo-angiogenesis.
Available evidence obtained from the different models studied
so far indicates that constitutive activation of NFAT through
calcineurin-dependent or independent pathways clearly contrib-
utes to the expression of one or more of the phenotypic traits that
characterize in vitro transformed- and tumor cells. First, enforced
expression in the 3T3L1 pre-adipocyte cell line of a constitutively
nuclear and transcriptionally active NFATc1 mutant (caNFATc1)
obtained by substitution of the phospshorylation/dephosphoryla-
tion sites in NFATc1 regulatory domain by alanine, was sufficient to
impair terminal differentiation into adipocytes and to induce cellular
transformation.
26
The bypass of the G
1
cell cycle checkpoint and
long-term proliferation induced by caNFATc1 under reduced serum
conditions was associated with upregulation of c-MYC, cyclin D2
and cyclin D3 expression, but whether this reflects direct transcrip-
tional deregulation of these genes by NFATc1 was not determined.
Enforced expression of caNFATc1 also protected cells from apoptosis
normally induced in 3T3-L1 cells in response to complete serum
withdrawal, an effect that was associated with the production of
autocrine/paracrine survival factors by transformed cells.
26
In cell
lines derived from pancreatic adenocarcinoma, CsA treatment and
siRNA-mediated knockdown of NFATc1 inhibited their prolif-
eration and anchorage-independent growth.
14
Cell lines that resisted
CsA treatment were derived from tumors harboring an amplification
of the c-MYC protooncogene whereas NFATc1-dependent regula-
tion of c-MYC was observed in responsive cells. An NFATc1 binding
site was identified in the c-MYC promoter, that overlaps with a
previously identified TGF-response element. Promoter studies in
transient transfection assays showed NFATc1-mediated activation
of a c-MYC promoter construct, suggesting the direct deregula-
tion of c-MYC expression by overexpressed NFATc1 in pancreatic
cancer.
14
Importantly, enforced expression of c-MYC was found
to rescue Panc1 cells proliferation from CsA-mediated calcineurin
inhibition, indicating that c-MYC is indeed a downstream target
of calcineurin/NFATc1 activation in these cells. In DLBCL-derived
cell lines, Ford and colleagues provided evidence that siRNA-medi-
ated downregulation of NFATc1 expression results in decreased cell
proliferation, impaired cell survival as well as reduced expression of
CD154, a ligand of the TNF family that binds CD40. Since DLBCL
express CD40, assembly of an autocrine stimulatory mechanism,
resulting in the assembly of a CD40 signalososme is proposed to
control cell survival and proliferation of DLBCL cell lines.
33
NFAT
binding sites were identified in the CD154 promoter and shown to
be important for promoter activity in DLBCL cell lines, a property
that relied upon NFATc1 acting in synergy with specific members of
the NFB family.
16
In the breast carcinoma cell line model, enforced
expression of NFATc2 was found to be sufficient to enhance cell
invasion in-matrigel assays whereas overexpression of either NFATc2
or NFAT5 induced cell migration.
13
Further studies have shown
that NFATc2 overexpression promotes the in vitro invasive pheno-
type of breast carcinoma cell lines through the induction of genes
such as COX2 (cyclooxygenase 2) and autotaxin in breast carcinoma
cells.
34,35
NFATc2 binding sites are found in the promoters of
both the autotaxin and COX2 genes, but whether NFATc2 directly
deregulates transcription of these genes in breast carcinome cells was
not analyzed.
In line with their activity as either positive or negative regulators of
genes involved in cell survival and proliferation, NFAT proteins have
also been described as potential tumor suppressor genes in specific
cellular contexts. Ectopic expression of NFATc2 has been reported to
promote apoptosis of Burkitt lymphoma derived cell lines, presum-
ably through the induction of Nur77, a member of the orphan
nuclear receptor superfamily, a transcriptional regulator involved
in apoptosis.
36
In addition, NFATc3-deficient mice infected by the
murine lymphomagenic retrovirus SL3-3 develop T-cell lymphomas
faster and with higher frequencies as compared to wild-type mice or
NFATc2-deficient mice,
37
suggesting that NFATc3 could act as a
tumor suppressor gene in the T cell lineage.
www.landesbioscience.com Cell Cycle 299
The calcineurin/NFAT signaling pathway in cancer
Available evidence thus indicates that, depending upon the tumor
type considered, an array of deregulated target genes may be acting
downstream of activated calcineurin/NFAT to modulate the tumor
phenotype. However, a number of questions remain to be answered.
First, the in vivo relevance of these observations, mostly made in cell
lines, should be addressed in appropriate mouse models of the respec-
tive cancers. Second, several NFAT factors are likely to be expressed
in tumor cells and the question arises whether the different NFAT
play a redundant or specific role in tumorigenesis. For example, clear
evidence indicates that different NFATs can have either specific, or
redundant or even antagonistic roles in diverse aspects of normal T
cell development (reviewed in ref. 8). This complexity also exists in
other cell lineages but to what extent its perturbation contributes to
tumor development at different phases of tumorigenesis is at present
unknown and will likely be distinct in different cancer types.
Finally, although mouse genetic studies have demonstrated a
strong epistatic relationship between calcineurin and NFATs activa-
tion, it is important to note that these transcription factors are not
the only calcineurin downstream substrates. Indeed, calcineurin has
been shown to positively regulate a number of other targets such as:
channels like InsP3R, the ryanodine or the NMDA receptors;
38,39
enzymes like PKA,
40
NO synthase;
41
other transcription factors like
MEF2C,
42
MEF2D
43
and the co-activator TORC2;
44
mechanisms
involved in mRNA stabilization
45
or miRNA expression.
46
Several
of these downstream targets could potentially mediate calcineurin
effects in tumor cells.
Upstream Signals Leading to Calcineurin/NFAT Activation
in Cancer Cells
Little information is available concerning the nature of the
upstream events that control calcineurin and NFAT activation in
tumor cells. Calcineurin-independent upregulation of NFATc2 and
NFAT5 expression in mammary tumor cell lines is linked to expres-
sion of 4 integrin and the assembly of an 64 heterodimer, but the
molecular events downstream of 64 have not been identified.
13
In
the TEL-JAK2 and ICN1-induced mouse models we have used,
17
the
rapid emergence of leukemia is critically dependent upon pre-TCR
signaling and these clonal leukemias express a functional TCR.
47,48
Since the pre-TCR and the TCR are well characterized receptors
coupled to calcium/calcineurin/NFAT activation,
9,49
we consid-
ered the possibility that sustained calcium/calcineurin signaling in
leukemic cells could reflect acquired hypersensitivity of these cells
to pre-TCR/TCR-dependent signals. This is not the case since both
TEL-JAK2- and ICN1-induced leukemias generated in Rag2-defi-
cient micewhich cannot rearrange the genes encoding TCR and
and thus cannot express neither the pre-TCR nor a TCRpresent
persistent activation of the calcineurin/NFAT signaling module in
a similar fashion as leukemias obtained in RAG2-proficient mice
(ref. 17 and our unpublished data). In our models of TEL-JAK2
and ICN1-induced T-ALL/lymphoma, continuous activation of the
calcineurin/NFAT module was interrupted when leukemic cells were
removed from their in vivo environment (e.g., thymus, spleen, lymph
nodes) and maintained in tissue culture for short periods of time.
17
This in vitro inactivation is reversible as re-implantation of these cells
into syngeneic hosts resulted in reactivation of the calcineurin/NFAT
pathway in the transplanted leukemias (our unpublished observa-
tions). This suggests that in vivo activation of the calcineurin/NFAT
pathway in leukemic cells depends upon autocrine/paracrine signals
or signals generated by other cells in the tumor micro-environ-
ment. This in vivo/in vitro dichotomy was found in other instances,
including human lymphoma samples (our unpublished observa-
tions). This also suggests that activation of the calcineurin/NFAT
pathway in lymphoid malignancies might be broader than previously
anticipated.
15-17
Our experiments also point to the fact that the TEL-JAK2 and
ICN1 initiating/maintenance oncoproteins are not sufficient to acti-
vate the calcineurin/NFAT module as they remained active under
in vitro conditions.
17
This does not exclude that they nevertheless
could assist other upstream events in calcineurin/NFAT activation.
For example, TEL-JAK2 is known to activate the endogenous AKT
protein kinase
50
and persistent activation of AKT could lead to
inactivation of GSK3, an export NFAT kinase. Likewise, in kera-
tinocytes, ICN1 has been shown to induce the Hes1-dependent
repression of the gene encoding calcipressin1 (CSP1), an endogenous
inhibitor of calcineurin.
51
Whether this regulation occurs in T-ALL
carrying activating mutations in NOTCH1
52
or in other malignan-
cies or other cancers remains to be investigated.
Cell lines derived from DLBCL and a subset of T-ALL cell lines
show activation of the calcineurin/NFAT module,
16,17
suggesting
that their establishment selected clones carrying mutations leading
to activation of this pathway. Such mutations have in fact been
described in lymphoma-derived cell lines. For example, the EL4
mouse T cell lymphoma cell line was found to express a mutant form
of calcineurin in which negative regulation of phosphatase activity
by the CnA autoinhibitory domain is impaired due to a mutation
that replaces an aspartic acid in position 477 by asparagine.
53
In
contrast, constitutive activation of calcineurin in the SML B-cell
lymphoma cell line was found to result from the expression of a
truncated version of the CnA catalytic subunit.
54
Proteolytic activa-
tion of caspases appears to be an alternative mode of activation of
calcineurin. To date, two different proteases, namely caspase3 and m-
calpain, have been shown to induce calcineurin cleavage in a way that
enhances its phosphatase activity.
55-57
In Jurkat T cells, activation of
caspase3 in response to PHA treatment leads to the cleavage of CnA
to generate truncated polypeptides with enhanced catalytic activity.
55
The second protease, m-calpain, activates calcineurin through two
distinct mechanisms: (i) the cleavage of CnA that removes its auto-
inhibiory domain, (ii) the cleavage of CABIN1, an endogenous
inhibitor of calcineurin. However, no cleaved forms of CnA were
detected in our T-ALL mouse models in which we have shown that
persistent calcineurin activation contributes to the leukemogenic
process (our unpublished observations).
Mutations in other components of calcium/calmodulin signaling
could be involved in the deregulation of calcineurin or other path-
ways in tumor cells. Under physiological conditions, calcineurin
activity is negatively regulated by a number of endogenous proteins
including the calcipressin family (CSP1, CSP2 and CSP3),
58
CABIN1 (CAIN),
59
AKAP79 (KAP5),
60,61
CHP (Calcineurin
Homologous Protein)
62
and FKBP38.
63
Therefore, dysregulated
expression of these negative regulators could contribute to enhanced
calcineurin activation observed in tumor cells. It has been shown that
T cells from mouse expressing a truncated version of CABIN1, that
is no longer capable of inhibiting calcineurin activity, overexpress
a number of cytokine genes (IL2, IL4, IL9, IL13, IFN) due to
300 Cell Cycle 2008; Vol. 7 Issue 3
The calcineurin/NFAT signaling pathway in cancer
their hypersensitivity to TCR signals
64
whereas downregulation of
DSCR1 favors the anchorage-dependent and anchorage-independent
proliferation of a colorectal cancer-derived cell line.
65
Persistent
calcineurin activation could also potentially result from deregulated
activity of upstream components in calcium signaling. For example,
the TRPV6 calcium channel is overexpressed in advanced stages of
prostate cancer and studies in the LNCaP prostate cancer cell line
show that TRPV6-mediated calcium import is associated with NFAT
activation and favors cell survival and proliferation.
66
Calcineurin/NFAT Signaling Pathway as a Potential Target
for Therapy
The growing body of evidence implicating the activation of the
calcineurin/NFAT signaling pathway in progression and/or mainte-
nance of solid tumors, lymphoma and leukemia suggests that the use
or development of inhibitors of the molecular events acting upstream
of calcineurin activation, of calcineurin itself or of critical effectors of
calcineurin may be useful in the treatment of these pathologies. CsA
and FK506 are structurally unrelated, well characterized immuno-
suppressive agents that function as co-drugs after binding to specific
endogenous cytoplasmic cyclophilins, namely cyclophilinA (CypA)
and FKB12, respectively. Both the CsA-CypA and FK506-FKBP12
complexes physically interact with calcineurin to inhibit its ability to
dephosphorylate protein substrates. This property is at the heart of
the potent inhibitory properties of CsA and FK506 in the response
of T cells to alloantigens and thus of the widespread clinical use of
CsA and FK506 to prevent the rejection of organ transplants and
in the treatment of aggressive forms of rheumatoid arthritis and
psoriasis.
7,69
T-ALL accounts for about 15% of pediatric and 25% of adult cases
of ALL. Molecular characterization of T-ALL has identified a number
of genes that contribute to cell cycle and growth deregulation (e.g., loss
of the CDKN2A locus), impaired differentiation (e.g., deregulated
expression of specific Hox genes) and unlimited self-renewal capacity
of leukemic cells. Irrespective of their stage of differentiation arrest,
approximately 5060% of T-ALL harbor activating mutations in the
NOTCH1 gene,
52
implying a central role of deregulated NOTCH
signaling in several aspects of T-ALL biology. While 75% of children
with T-cell ALL are cured with combination chemotherapy, the
remaining 25% suffer either from refractory or relapsed diseases. These
figures and the toxic side effects associated with available chemotherapy
regimens clearly calls for the search of novel therapeutic options. Our
studies in several mouse models of human T-cell leukemia, including
the T-ALL/lymphoma induced by activated NOTCH1, show that
short-term (714 days) treatment with either CsA or FK506 results
in clear anti-leukemic effects and in prolonged survival of treated
versus non treated animals.
15,17
These pre-clinical data suggests that
the use of these compounds could provide a therapeutic benefit in
remission induction or in the consolidation phase of T-ALL treatment
and possibly other malignancies that display high calcineurin/NFAT
activation.
13,14,16,32
However CsA and FK506 show severe toxic side
effects (neurotoxicity, nephrotoxicity, gastrointestinal disturbances,
hypertension) and their long term administration (10 years on longer)
in transplanted patients is associated with the emergence of specific
cancers due to suppression of tumor immunosurveillance mechanisms.
These side effects may compromise the usefulness of CsA and FK506
even in short term induction remission protocols. Nevertheless, case
reports have described long term remission of specific lymphocytic
leukemias after CsA treatment
67
and the additional inhibitory prop-
erties of CsA on ABC transporters have been proposed to form the
basis of its therapeutic benefit in AML.
68
Interestingly, derivatives of
CsA and FK506 have recently been described, including L-732531
(an analog of FK506
69
) and ISATX247 (an analog of CsA
70,71
), that
show comparable or even higher efficiency toward calcineurin inhibi-
tion and reduced renal toxicity as compared to the respective lead
compound.
69-71
The alledged limitations linked to direct inhibition of calcineurin
activity clearly requires the further dissection of the molecular path-
ways involved in its activation in human malignancies. To date,
the molecular events leading to persistent calcineurin activation in
cancers remain to be identified and, as discussed above are likely to
be distinct in different cancer types. Prior studies have clearly demon-
strated that under physiological conditions, calcineurin activation is
dependent upon the increase in intracellular calcium concentration
through a capacitative calcium entry (CCE) via the CRAC chan-
nels (reviewed in ref. 7 and references herein). Therefore, it might
be expected that inhibition of CCE through specific inhibitors
of the CRAC channels would have similar effects as inhibition of
calcineurin with CsA and FK506 (reviewed in ref. 7). Two different
compounds, namely BTP2 and capsaicin, which fulfill these criteria
have recently been described
72-74
and therefore could represent
an alternative approach to inhibit calcineurin for the treatment of
cancers where persistent calcineurin activation is implicated. These
therapeutic options however are likely to suffer from the same limita-
tions as those linked to the direct inhibition of calcineurin.
It is therefore critical to identify and analyze the respective roles
of the downstream effectors involved in the pro-oncogenic activity
of calcineurin in different malignancies. As discussed above, available
data support the implication of the NFAT family of transcrip-
tion factors in cancer and the relevant genes/pathways deregulated
by constitutive NFAT activation may represent novel, potential
therapeutic targets. However, some of the other previously described
calcineurin targets have also been implicated in cancer. For example,
the MEF2D transcription factor is a member of the MEF2 family of
DNA binding proteins that activate transcription of genes involved
in the control of muscle cell differentiation and in the response of
neuronal cells and T-lymphocytes to mitogenic and survival signals.
In murine retroviral insertional mutagenesis studies, MEF2D has
been identified as a candidate oncogene involved in the pathogen-
esis of leukemia.
75,76
Thus, deregulation of MEF2 target genes, if it
occurs in tumor cells harboring persistent calcineurin activation, may
offer alternative targets of therapeutic interest.
Acknowledgements
HM was supported by fellowships from the Ministre de
lEducation Nationale et de la Recherche and lAssociation pour la
Recherche contre le Cancer (ARC). Our work was supported by
funds from CNRS (Centre National de la Recherche Scientifique),
Institut Curie, INCA (Institut National du Cancer), Cancrople
Ile-de-France, ANR (Agence Nationale de la Recherche), Ligue
Nationale contre le Cancer (quipe Labelise Ligue) and Association
for International Cancer Research.
www.landesbioscience.com Cell Cycle 301
The calcineurin/NFAT signaling pathway in cancer
References
1. Hogan PG, Chen L, Nardone J, Rao A. Transcriptional regulation by calcium, calcineurin,
and NFAT. Genes & development 2003; 17:2205-32.
2. Klee CB, Ren H, Wang X. Regulation of the calmodulin-stimulated protein phosphatase,
calcineurin. The Journal of biological chemistry 1998; 273:13367-70.
3. Wu H, Peisley A, Graef IA, Crabtree GR. NFAT signaling and the invention of vertebrates.
Trends in cell biology 2007; 17:251-60.
4. Rusnak F, Mertz P. Calcineurin: form and function. Physiological reviews 2000;
80:1483-521.
5. Crabtree GR. Generic signals and specific outcomes: signaling through Ca2+, calcineurin,
and NF-AT. Cell 1999; 96:611-4.
6. Graef IA, Chen F, Crabtree GR. NFAT signaling in vertebrate development. Current opin-
ion in genetics & development 2001; 11:505-12.
7. Gwack Y, Feske S, Srikanth S, Hogan PG, Rao A. Signalling to transcription: store-operated
Ca2+ entry and NFAT activation in lymphocytes. Cell calcium 2007; 42:145-56.
8. Macian F. NFAT proteins: key regulators of T-cell development and function. Nature
reviews 2005; 5:472-84.
9. Neilson JR, Winslow MM, Hur EM, Crabtree GR. Calcineurin B1 is essential for positive
but not negative selection during thymocyte development. Immunity 2004; 20:255-66.
10. Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, Tanasa B, Hogan PG, Lewis RS,
Daly M, Rao A. A mutation in Orai1 causes immune deficiency by abrogating CRAC chan-
nel function. Nature 2006; 441:179-85.
11. Heit JJ, Apelqvist AA, Gu X, Winslow MM, Neilson JR, Crabtree GR, Kim SK.
Calcineurin/NFAT signalling regulates pancreatic beta-cell growth and function. Nature
2006; 443:345-9.
12. Buchholz M, Ellenrieder V. An emerging role for Ca2+/calcineurin/NFAT signaling in
cancerogenesis. Cell cycle (Georgetown, Tex 2007; 6:16-9.
13. Jauliac S, Lopez-Rodriguez C, Shaw LM, Brown LF, Rao A, Toker A. The role of NFAT
transcription factors in integrin-mediated carcinoma invasion. Nature cell biology 2002;
4:540-4.
14. Buchholz M, Schatz A, Wagner M, Michl P, Linhart T, Adler G, Gress TM, Ellenrieder V.
Overexpression of c-myc in pancreatic cancer caused by ectopic activation of NFATc1 and
the Ca2+/calcineurin signaling pathway. The EMBO journal 2006; 25:3714-24.
15. Marafioti T, Pozzobon M, Hansmann ML, Ventura R, Pileri SA, Roberton H, Gesk S,
Gaulard P, Barth TF, Du MQ, Leoncini L, Moller P, Natkunam Y, Siebert R, Mason DY.
The NFATc1 transcription factor is widely expressed in white cells and translocates from the
cytoplasm to the nucleus in a subset of human lymphomas. British journal of haematology
2005; 128:333-42.
16. Pham LV, Tamayo AT, Yoshimura LC, Lin-Lee YC, Ford RJ. Constitutive NFkappaB and
NFAT activation in aggressive B-cell lymphomas synergistically activates the CD154 gene
and maintains lymphoma cell survival. Blood 2005; 106:3940-7.
17. Medyouf H, Alcalde H, Berthier C, Guillemin MC, dos Santos NR, Janin A, Decaudin D,
de The H, Ghysdael J. Targeting calcineurin activation as a therapeutic strategy for T-cell
acute lymphoblastic leukemia. Nature medicine 2007; 13:736-41.
18. Clipstone NA, Crabtree GR. Identification of calcineurin as a key signalling enzyme in T-
lymphocyte activation. Nature 1992; 357:695-7.
19. Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR. Identification of a puta-
tive regulator of early T cell activation genes. Science (New York, NY 1988; 241:202-5.
20. Kahl CR, Means AR. Regulation of cell cycle progression by calcium/calmodulin-dependent
pathways. Endocrine reviews 2003; 24:719-36.
21. Baksh S, DeCaprio JA, Burakoff SJ. Calcineurin regulation of the mammalian G
0
/G
1
checkpoint element, cyclin dependent kinase 4. Oncogene 2000; 19:2820-7.
22. Baksh S, Widlund HR, Frazer-Abel AA, Du J, Fosmire S, Fisher DE, DeCaprio JA,
Modiano JF, Burakoff SJ. NFATc2-mediated repression of cyclin-dependent kinase 4
expression. Molecular cell 2002; 10:1071-81.
23. Carvalho LD, Teixeira LK, Carrossini N, Caldeira AT, Ansel KM, Rao A, Viola JP.
The NFAT1 transcription factor is a repressor of cyclin A2 gene expression. Cell cycle
(Georgetown, Tex 2007; 6:1789-95.
24. Santini MP, Talora C, Seki T, Bolgan L, Dotto GP. Cross talk among calcineurin, Sp1/Sp3,
and NFAT in control of p21(WAF1/CIP1) expression in keratinocyte differentiation.
Proceedings of the National Academy of Sciences of the United States of America 2001;
98:9575-80.
25. Caetano MS, Vieira-de-Abreu A, Teixeira LK, Werneck MB, Barcinski MA, Viola JP.
NFATC2 transcription factor regulates cell cycle progression during lymphocyte activa-
tion: evidence of its involvement in the control of cyclin gene expression. Faseb J 2002;
16:1940-2.
26. Neal JW, Clipstone NA. A constitutively active NFATc1 mutant induces a transformed phe-
notype in 3T3-L1 fibroblasts. The Journal of biological chemistry 2003; 278:17246-54.
27. Oukka M, Ho IC, de la Brousse FC, Hoey T, Grusby MJ, Glimcher LH. The transcrip-
tion factor NFAT4 is involved in the generation and survival of T cells. Immunity 1998;
9:295-304.
28. Cante-Barrett K, Winslow MM, Crabtree GR. Selective role of NFATc3 in positive selec-
tion of thymocytes. J Immunol 2007; 179:103-10.
29. Graef IA, Wang F, Charron F, Chen L, Neilson J, Tessier-Lavigne M, Crabtree GR.
Neurotrophins and netrins require calcineurin/NFAT signaling to stimulate outgrowth of
embryonic axons. Cell 2003; 113:657-70.
30. Hernandez GL, Volpert OV, Iniguez MA, Lorenzo E, Martinez-Martinez S, Grau R, Fresno
M, Redondo JM. Selective inhibition of vascular endothelial growth factor-mediated angio-
genesis by cyclosporin A: roles of the nuclear factor of activated T cells and cyclooxygenase
2. The Journal of experimental medicine 2001; 193:607-20.
31. Graef IA, Chen F, Chen L, Kuo A, Crabtree GR. Signals transduced by Ca(2+)/calcineurin
and NFATc3/c4 pattern the developing vasculature. Cell 2001; 105:863-75.
32. Zaichuk TA, Shroff EH, Emmanuel R, Filleur S, Nelius T, Volpert OV. Nuclear factor of
activated T cells balances angiogenesis activation and inhibition. The Journal of experimen-
tal medicine 2004; 199:1513-22.
33. Pham LV, Tamayo AT, Yoshimura LC, Lo P, Terry N, Reid PS, Ford RJ. A CD40
Signalosome anchored in lipid rafts leads to constitutive activation of NFkappaB and
autonomous cell growth in B cell lymphomas. Immunity 2002; 16:37-50.
34. Chen M, OConnor KL. Integrin alpha6beta4 promotes expression of autotaxin/ENPP2
autocrine motility factor in breast carcinoma cells. Oncogene 2005; 24:5125-30.
35. Yiu GK, Toker A. NFAT induces breast cancer cell invasion by promoting the induction of
cyclooxygenase-2. The Journal of biological chemistry 2006; 281:12210-7.
36. Kondo E, Harashima A, Takabatake T, Takahashi H, Matsuo Y, Yoshino T, Orita K, Akagi
T. NF-ATc2 induces apoptosis in Burkitts lymphoma cells through signaling via the B cell
antigen receptor. European journal of immunology 2003; 33:1-11.
37. Glud SZ, Sorensen AB, Andrulis M, Wang B, Kondo E, Jessen R, Krenacs L, Stelkovics
E, Wabl M, Serfling E, Palmetshofer A, Pedersen FS. A tumor-suppressor function for
NFATc3 in T-cell lymphomagenesis by murine leukemia virus. Blood 2005; 106:3546-52.
38. Cameron AM, Steiner JP, Roskams AJ, Ali SM, Ronnett GV, Snyder SH. Calcineurin
associated with the inositol 1,4,5-trisphosphate receptor-FKBP12 complex modulates Ca2+
flux. Cell 1995; 83:463-72.
39. Tong G, Shepherd D, Jahr CE. Synaptic desensitization of NMDA receptors by calcineurin.
Science (New York, NY 1995; 267:1510-2.
40. Blumenthal DK, Takio K, Hansen RS, Krebs EG. Dephosphorylation of cAMP-depen-
dent protein kinase regulatory subunit (type II) by calmodulin-dependent protein phos-
phatase. Determinants of substrate specificity. The Journal of biological chemistry 1986;
261:8140-5.
41. Dawson VL, Dawson TM, Uhl GR, Snyder SH. Human immunodeficiency virus type 1
coat protein neurotoxicity mediated by nitric oxide in primary cortical cultures. Proceedings
of the National Academy of Sciences of the United States of America 1993; 90:3256-9.
42. Lynch J, Guo L, Gelebart P, Chilibeck K, Xu J, Molkentin JD, Agellon LB, Michalak M.
Calreticulin signals upstream of calcineurin and MEF2C in a critical Ca(2+)-dependent
signaling cascade. The Journal of cell biology 2005; 170:37-47.
43. Wu H, Rothermel B, Kanatous S, Rosenberg P, Naya FJ, Shelton JM, Hutcheson KA,
DiMaio JM, Olson EN, Bassel-Duby R, Williams RS. Activation of MEF2 by muscle
activity is mediated through a calcineurin-dependent pathway. The EMBO journal 2001;
20:6414-23.
44. Screaton RA, Conkright MD, Katoh Y, Best JL, Canettieri G, Jeffries S, Guzman E, Niessen
S, Yates JR, 3rd, Takemori H, Okamoto M, Montminy M. The CREB coactivator TORC2
functions as a calcium- and cAMP-sensitive coincidence detector. Cell 2004; 119:61-74.
45. Marshall RM, Salerno D, Garriga J, Grana X. Cyclin T1 expression is regulated by multiple
signaling pathways and mechanisms during activation of human peripheral blood lympho-
cytes. J Immunol 2005; 175:6402-11.
46. van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, Gerard RD, Richardson
JA, Olson EN. A signature pattern of stress-responsive microRNAs that can evoke cardiac
hypertrophy and heart failure. Proceedings of the National Academy of Sciences of the
United States of America 2006; 103:18255-60.
47. Campese AF, Garbe AI, Zhang F, Grassi F, Screpanti I, von Boehmer H. Notch1-dependent
lymphomagenesis is assisted by but does not essentially require pre-TCR signaling. Blood
2006; 108:305-10.
48. dos Santos NR, Rickman DS, de Reynies A, Cormier F, Williame M, Blanchard C, Stern
MH, Ghysdael J. Pre-TCR expression cooperates with TEL-JAK2 to transform immature
thymocytes and induce T-cell leukemia. Blood 2007; 109:3972-81.
49. Aifantis I, Gounari F, Scorrano L, Borowski C, von Boehmer H. Constitutive pre-TCR
signaling promotes differentiation through Ca2+ mobilization and activation of NFkappaB
and NFAT. Nature immunology 2001; 2:403-9.
50. Nguyen MH, Ho JM, Beattie BK, Barber DL. TEL-JAK2 mediates constitutive activation
of the phosphatidylinositol 3'-kinase/protein kinase B signaling pathway. The Journal of
biological chemistry 2001; 276:32704-13.
51. Mammucari C, Tommasi di Vignano A, Sharov AA, Neilson J, Havrda MC, Roop DR,
Botchkarev VA, Crabtree GR, Dotto GP. Integration of Notch 1 and calcineurin/NFAT
signaling pathways in keratinocyte growth and differentiation control. Developmental cell
2005; 8:665-76.
52. Weng AP, Ferrando AA, Lee W, Morris JPt, Silverman LB, Sanchez-Irizarry C, Blacklow SC,
Look AT, Aster JC. Activating mutations of NOTCH1 in human T cell acute lymphoblastic
leukemia. Science (New York, NY 2004; 306:269-71.
53. Fruman DA, Pai SY, Burakoff SJ, Bierer BE. Characterization of a mutant calcineurin
A alpha gene expressed by EL4 lymphoma cells. Molecular and cellular biology 1995;
15:3857-63.
54. Gross KL, Cioffi EA, Scammell JG. Increased activity of the calcineurin-nuclear factor of
activated T cells pathway in squirrel monkey B-Lymphoblasts identified by PowerBlot. In
vitro cellular & developmental biology 2004; 40:57-63.
302 Cell Cycle 2008; Vol. 7 Issue 3
The calcineurin/NFAT signaling pathway in cancer
55. Mukerjee N, McGinnis KM, Gnegy ME, Wang KK. Caspase-mediated calcineurin acti-
vation contributes to IL-2 release during T cell activation. Biochemical and biophysical
research communications 2001; 285:1192-9.
56. Kim MJ, Jo DG, Hong GS, Kim BJ, Lai M, Cho DH, Kim KW, Bandyopadhyay A, Hong
YM, Kim DH, Cho C, Liu JO, Snyder SH, Jung YK. Calpain-dependent cleavage of
cain/cabin1 activates calcineurin to mediate calcium-triggered cell death. Proceedings of the
National Academy of Sciences of the United States of America 2002; 99:9870-5.
57. Wu HY, Tomizawa K, Oda Y, Wei FY, Lu YF, Matsushita M, Li ST, Moriwaki A, Matsui H.
Critical role of calpain-mediated cleavage of calcineurin in excitotoxic neurodegeneration.
The Journal of biological chemistry 2004; 279:4929-40.
58. Kingsbury TJ, Cunningham KW. A conserved family of calcineurin regulators. Genes &
development 2000; 14:1595-604.
59. Sun L, Youn HD, Loh C, Stolow M, He W, Liu JO. Cabin 1, a negative regulator for
calcineurin signaling in T lymphocytes. Immunity 1998; 8:703-11.
60. Coghlan VM, Perrino BA, Howard M, Langeberg LK, Hicks JB, Gallatin WM, Scott JD.
Association of protein kinase A and protein phosphatase 2B with a common anchoring
protein. Science (New York, NY 1995; 267:108-11.
61. Klauck TM, Faux MC, Labudda K, Langeberg LK, Jaken S, Scott JD. Coordination of
three signaling enzymes by AKAP79, a mammalian scaffold protein. Science (New York,
NY 1996; 271:1589-92.
62. Lin X, Sikkink RA, Rusnak F, Barber DL. Inhibition of calcineurin phosphatase activ-
ity by a calcineurin B homologous protein. The Journal of biological chemistry 1999;
274:36125-31.
63. Shirane M, Nakayama KI. Inherent calcineurin inhibitor FKBP38 targets Bcl-2 to mito-
chondria and inhibits apoptosis. Nature cell biology 2003; 5:28-37.
64. Esau C, Boes M, Youn HD, Tatterson L, Liu JO, Chen J. Deletion of calcineurin and
myocyte enhancer factor 2 (MEF2) binding domain of Cabin1 results in enhanced cytokine
gene expression in T cells. The Journal of experimental medicine 2001; 194:1449-59.
65. Bush CR, Havens JM, Necela BM, Su W, Chen L, Yanagisawa M, Anastasiadis PZ, Guerra
R, Luxon BA, Thompson EA. Functional genomic analysis reveals cross-talk between per-
oxisome proliferator-activated receptor gamma and calcium signaling in human colorectal
cancer cells. The Journal of biological chemistry 2007; 282:23387-401.
66. Lehenkyi V, Flourakis M, Skryma R, Prevarskaya N. TRPV6 channel controls prostate
cancer cell proliferation via Ca(2+)/NFAT-dependent pathways. Oncogene 2007.
67. Gabor EP, Mishalani S, Lee S. Rapid response to cyclosporine therapy and sustained remis-
sion in large granular lymphocyte leukemia. Blood 1996; 87:1199-200.
68. List AF, Kopecky KJ, Willman CL, Head DR, Slovak ML, Douer D, Dakhil SR,
Appelbaum FR. Cyclosporine inhibition of P-glycoprotein in chronic myeloid leukemia
blast phase. Blood 2002; 100:1910-2.
69. Dumont FJ. FK506, an immunosuppressant targeting calcineurin function. Current
medicinal chemistry 2000; 7:731-48.
70. Aspeslet L, Freitag D, Trepanier D, Abel M, Naicker S, Kneteman N, Foster R, Yatscoff R.
ISA(TX)247: a novel calcineurin inhibitor. Transplantation proceedings 2001; 33:1048-51.
71. Stalder M, Birsan T, Hubble RW, Paniagua RT, Morris RE. In vivo evaluation of the novel
calcineurin inhibitor ISATX247 in non-human primates. J Heart Lung Transplant 2003;
22:1343-52.
72. Ishikawa J, Ohga K, Yoshino T, Takezawa R, Ichikawa A, Kubota H, Yamada T. A pyrazole
derivative, YM-58483, potently inhibits store-operated sustained Ca2+ influx and IL-2
production in T lymphocytes. J Immunol 2003; 170:4441-9.
73. Zitt C, Strauss B, Schwarz EC, Spaeth N, Rast G, Hatzelmann A, Hoth M. Potent inhibi-
tion of Ca2+ release-activated Ca2+ channels and T-lymphocyte activation by the pyrazole
derivative BTP2. The Journal of biological chemistry 2004; 279:12427-37.
74. Fischer BS, Qin D, Kim K, McDonald TV. Capsaicin inhibits Jurkat T-cell activation by
blocking calcium entry current I(CRAC). The Journal of pharmacology and experimental
therapeutics 2001; 299:238-46.
75. Lund AH, Turner G, Trubetskoy A, Verhoeven E, Wientjens E, Hulsman D, Russell R,
DePinho RA, Lenz J, van Lohuizen M. Genome-wide retroviral insertional tagging of genes
involved in cancer in Cdkn2a-deficient mice. Nature genetics 2002; 32:160-5.
76. Suzuki T, Shen H, Akagi K, Morse HC, Malley JD, Naiman DQ, Jenkins NA, Copeland
NG. New genes involved in cancer identified by retroviral tagging. Nature genetics 2002;
32:166-74.
www.landesbioscience.com Cell Cycle 303
Você também pode gostar
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Mendelian Genetics - PPT Iiird SemDocumento70 páginasMendelian Genetics - PPT Iiird SemMousumi DasAinda não há avaliações
- Chapter 9 - Plant Biology: MarkschemeDocumento17 páginasChapter 9 - Plant Biology: MarkschemeAtulya BharadwajAinda não há avaliações
- Brainpower: Complex Organ Controls Your Every Thought and MoveDocumento13 páginasBrainpower: Complex Organ Controls Your Every Thought and MoveEve Laurence Nicolas100% (4)
- Soojeedes Human Anatomy and Physiology PDFDocumento119 páginasSoojeedes Human Anatomy and Physiology PDFmaxamed Buule IsaaqAinda não há avaliações
- Uncovering Principles of Brain EvolutionDocumento36 páginasUncovering Principles of Brain EvolutionCulture HeritageAinda não há avaliações
- Drug Safety AssessmentDocumento7 páginasDrug Safety AssessmentSAN912Ainda não há avaliações
- Current Developments in Activity-Based Protein Pro FilingDocumento11 páginasCurrent Developments in Activity-Based Protein Pro FilingSAN912Ainda não há avaliações
- HPLC Maintenance AgilentDocumento4 páginasHPLC Maintenance AgilentSAN912Ainda não há avaliações
- Guidance For Industry Estimating The Maximum Safe Starting Dose in Initial Clinical Trials For Therapeutics in Adult Healthy VolunteersDocumento30 páginasGuidance For Industry Estimating The Maximum Safe Starting Dose in Initial Clinical Trials For Therapeutics in Adult Healthy Volunteersaji_koeandaAinda não há avaliações
- The Science of Scientific Writing American ScientistDocumento7 páginasThe Science of Scientific Writing American ScientistSAN912Ainda não há avaliações
- Nicotinic ReceptorsDocumento13 páginasNicotinic ReceptorsSAN912Ainda não há avaliações
- Radhika ArticleDocumento27 páginasRadhika ArticleSAN912Ainda não há avaliações
- Us 6078908Documento10 páginasUs 6078908SAN912Ainda não há avaliações
- NFAT - Sp1 Cross TalkDocumento6 páginasNFAT - Sp1 Cross TalkSAN912Ainda não há avaliações
- United States Patent: Pathi Et A1. (10) Patent N0.: (45) Date of PatentDocumento8 páginasUnited States Patent: Pathi Et A1. (10) Patent N0.: (45) Date of PatentSAN912Ainda não há avaliações
- Paul PHD ThesisDocumento164 páginasPaul PHD ThesisSAN912Ainda não há avaliações
- Us 6078908Documento10 páginasUs 6078908SAN912Ainda não há avaliações
- Microsome S9 Prep ProtocolDocumento22 páginasMicrosome S9 Prep ProtocolSAN912Ainda não há avaliações
- Phoenix 1.3 Getting Started GuideDocumento70 páginasPhoenix 1.3 Getting Started GuideSAN912Ainda não há avaliações
- Phoenix 1.3 Getting Started GuideDocumento70 páginasPhoenix 1.3 Getting Started GuideSAN912Ainda não há avaliações
- Us 8557835Documento10 páginasUs 8557835SAN912Ainda não há avaliações
- United States: (12) Patent Application Publication (10) Pub. No.: US 2012/0270933 A1Documento18 páginasUnited States: (12) Patent Application Publication (10) Pub. No.: US 2012/0270933 A1SAN912Ainda não há avaliações
- Us 8557835Documento10 páginasUs 8557835SAN912Ainda não há avaliações
- Photosynthesis PDFDocumento30 páginasPhotosynthesis PDFhumble happyAinda não há avaliações
- Regulation of Gene ExpressionDocumento1 páginaRegulation of Gene ExpressionhomamunfatAinda não há avaliações
- Biology Class 8 Worksheet Reproduction in Plants and Animals, Growth and DevelopmentDocumento2 páginasBiology Class 8 Worksheet Reproduction in Plants and Animals, Growth and DevelopmentMEDHA mohanAinda não há avaliações
- 1 15 AstrovirusDocumento16 páginas1 15 AstrovirusRizal Hery100% (1)
- Bio ProjectDocumento17 páginasBio ProjectSpectra DragneelAinda não há avaliações
- Chapter 9 Inhibiting Growth of Pathogens in Vivo Using Antimicrobial AgentsDocumento5 páginasChapter 9 Inhibiting Growth of Pathogens in Vivo Using Antimicrobial AgentsMa. Kate Nicole D. PadpadAinda não há avaliações
- Ecosystem Structure and Types EcosystemDocumento27 páginasEcosystem Structure and Types EcosystemMOVIES starAinda não há avaliações
- Different Types of FermentationDocumento26 páginasDifferent Types of FermentationCats and DogAinda não há avaliações
- Alfalfa GrowthDocumento22 páginasAlfalfa GrowthEkaterina BogatyrevaAinda não há avaliações
- CARBOHYDRATE ABSORPTION (Ref)Documento22 páginasCARBOHYDRATE ABSORPTION (Ref)NEERAJA S RAJAinda não há avaliações
- Are We Prisoners of Our Genes?Documento18 páginasAre We Prisoners of Our Genes?pfbcarlisleAinda não há avaliações
- Questions & Answers: For NEET (UG) - 2020Documento22 páginasQuestions & Answers: For NEET (UG) - 2020Ashok Kumar PatelAinda não há avaliações
- 9.4. CellDocumento88 páginas9.4. CellTUĞRUL ERGÜLAinda não há avaliações
- Peripheral and Central Giant Cell Lesions: Etiology, Diagnosis and TreatmentDocumento7 páginasPeripheral and Central Giant Cell Lesions: Etiology, Diagnosis and TreatmentYeni PuspitasariAinda não há avaliações
- NIH Public AccessDocumento15 páginasNIH Public AccessAjaykumarAinda não há avaliações
- New Insights in Dialysis Membrane BiocompatibilityDocumento7 páginasNew Insights in Dialysis Membrane BiocompatibilityDani ursAinda não há avaliações
- Q1 Q2 Module 6 Cell CycleDocumento2 páginasQ1 Q2 Module 6 Cell Cycleindaljerson72Ainda não há avaliações
- Anaplastic meningioma diagnosisDocumento1 páginaAnaplastic meningioma diagnosisBhavik ChavdaAinda não há avaliações
- Toxoplasmosis: Zoonotic Disease Caused by Protozoan Parasite T. gondiiDocumento38 páginasToxoplasmosis: Zoonotic Disease Caused by Protozoan Parasite T. gondiiemmaAinda não há avaliações
- COVID Thrombosis: Mitochondria, The Endothelial Glycocalyx and Platelets Are KeyDocumento10 páginasCOVID Thrombosis: Mitochondria, The Endothelial Glycocalyx and Platelets Are KeyMercedes BouterAinda não há avaliações
- Act 2. Physiology Lab. (Yeo's Part)Documento3 páginasAct 2. Physiology Lab. (Yeo's Part)YEO, REGGIE ALBERT A.Ainda não há avaliações
- Advanced NanoBiomed Research - 2021 - Ratri - Recent Advances in Regenerative Tissue Fabrication Tools Materials andDocumento20 páginasAdvanced NanoBiomed Research - 2021 - Ratri - Recent Advances in Regenerative Tissue Fabrication Tools Materials andKalaaji WirasataAinda não há avaliações
- Presentation On Fungi SymbiosisDocumento19 páginasPresentation On Fungi SymbiosisFemi GajiAinda não há avaliações
- Cushnie and Lamb (2005)Documento16 páginasCushnie and Lamb (2005)ayu alfiantiAinda não há avaliações