Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Cruzeiro Pesticide Ape 2018Documento22 páginasCruzeiro Pesticide Ape 2018Popescu DragosAinda não há avaliações
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Alatorre Acvacultura Water QualityDocumento17 páginasAlatorre Acvacultura Water QualityPopescu DragosAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- HPLC Metoda de Validare PDFDocumento18 páginasHPLC Metoda de Validare PDFDiane DianaAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Agarwal OCP Rainwater 1986Documento10 páginasAgarwal OCP Rainwater 1986Popescu DragosAinda não há avaliações
- Barlas OCP Ape Turcia 2002Documento7 páginasBarlas OCP Ape Turcia 2002Popescu DragosAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Zinevici Lac Snagov 2014Documento14 páginasZinevici Lac Snagov 2014Popescu DragosAinda não há avaliações
- Pop Balea Lac 2012 PDFDocumento6 páginasPop Balea Lac 2012 PDFPopescu DragosAinda não há avaliações
- Vitamina C FructeDocumento5 páginasVitamina C FructePopescu DragosAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Adejo 2015Documento7 páginasAdejo 2015Popescu DragosAinda não há avaliações
- Importance of Sulphur in Crop ProductionDocumento18 páginasImportance of Sulphur in Crop ProductionPopescu DragosAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Fruit Maturity and Quality of Selectively Picked Royal Gala' ApplesDocumento6 páginasFruit Maturity and Quality of Selectively Picked Royal Gala' ApplesPopescu DragosAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Importance of Sulphur in Crop ProductionDocumento18 páginasImportance of Sulphur in Crop ProductionPopescu DragosAinda não há avaliações
- Importance of Sulphur in Crop ProductionDocumento18 páginasImportance of Sulphur in Crop ProductionPopescu DragosAinda não há avaliações
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Indicatori Calitate Tomate 1Documento7 páginasIndicatori Calitate Tomate 1Popescu DragosAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Aciditate Titrabila TomateDocumento3 páginasAciditate Titrabila TomatePopescu DragosAinda não há avaliações
- Calitate Depozitare Kiwi PDFDocumento7 páginasCalitate Depozitare Kiwi PDFPopescu DragosAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Indicatori Calitate ArdeiDocumento9 páginasIndicatori Calitate ArdeiPopescu DragosAinda não há avaliações
- Bordean 2011Documento5 páginasBordean 2011Popescu DragosAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Indicatori Calitate TomateDocumento5 páginasIndicatori Calitate TomatePopescu DragosAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Mineral Content of Apples Stored in Refrigeration Conditions and Controlled Atmosphere Conditions Respectively For 6 MonthsDocumento4 páginasMineral Content of Apples Stored in Refrigeration Conditions and Controlled Atmosphere Conditions Respectively For 6 MonthsPopescu DragosAinda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Instructions For Use: Hokkaido University Collection of Scholarly and Academic Papers: HUSCAPDocumento10 páginasInstructions For Use: Hokkaido University Collection of Scholarly and Academic Papers: HUSCAPPopescu DragosAinda não há avaliações
- Cadar 2014Documento5 páginasCadar 2014Popescu DragosAinda não há avaliações
- ArticolDocumento26 páginasArticolPopescu DragosAinda não há avaliações
- Antioxidant Ulei MaslineDocumento12 páginasAntioxidant Ulei MaslinePopescu DragosAinda não há avaliações
- JCC ReviewDocumento27 páginasJCC ReviewPopescu DragosAinda não há avaliações
- Maize Nutrient Accumulation and Partitioning in Response To Plant Density and Nitrogen Rate: I. MacronutrientsDocumento13 páginasMaize Nutrient Accumulation and Partitioning in Response To Plant Density and Nitrogen Rate: I. MacronutrientsPopescu DragosAinda não há avaliações
- LWR NigeriaDocumento3 páginasLWR NigeriaPopescu DragosAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (120)
- LWR TurciaDocumento5 páginasLWR TurciaPopescu DragosAinda não há avaliações
- Bod Cod TocDocumento24 páginasBod Cod TocPopescu DragosAinda não há avaliações
- IUPAC Press Release 30dec2015Documento3 páginasIUPAC Press Release 30dec2015Popescu DragosAinda não há avaliações
- DoctrineDocumento1 páginaDoctrinevinay44Ainda não há avaliações
- Maternity Benefit-Employer's ObligationsDocumento20 páginasMaternity Benefit-Employer's ObligationsSonika BhatiAinda não há avaliações
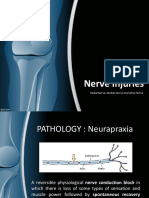
- Nerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveDocumento26 páginasNerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveaisiyaahAinda não há avaliações
- StoryDocumento6 páginasStoryGerard NicholasAinda não há avaliações
- Working in A Lab Risk & AssessmentDocumento10 páginasWorking in A Lab Risk & AssessmentMariam KhanAinda não há avaliações
- Affidavit - Rural Cess - ManjulaDocumento9 páginasAffidavit - Rural Cess - Manjulagebrsf setgwgvAinda não há avaliações
- Hazop PDFDocumento18 páginasHazop PDFLuiz Rubens Souza Cantelli0% (1)
- Research Paper About Poverty - NSTP Project For FINALSDocumento5 páginasResearch Paper About Poverty - NSTP Project For FINALSLeoAinda não há avaliações
- Therapy Is For Everybody: Black Mental HealthDocumento22 páginasTherapy Is For Everybody: Black Mental HealthIresha Picot100% (2)
- Foods SiddhasDocumento4 páginasFoods SiddhasSanjay JayaratneAinda não há avaliações
- A Review Article On Edible Pigments Properties and Sources As Natural Biocolorants in Foodstuff and Food IndustryDocumento8 páginasA Review Article On Edible Pigments Properties and Sources As Natural Biocolorants in Foodstuff and Food Industrybestread67Ainda não há avaliações
- The Varsity 39Documento254 páginasThe Varsity 39cosmin_bloju8997Ainda não há avaliações
- Philippine Science and Technology InnovationDocumento7 páginasPhilippine Science and Technology Innovationgabosara298Ainda não há avaliações
- YDA Nepal - MembersDocumento13 páginasYDA Nepal - MembersSulochan LohaniAinda não há avaliações
- Contentdamdiagnosticsusenproductsaaccu Chek Inform IitoolkitOS0130802 Accu Chek Inform IIDocumento264 páginasContentdamdiagnosticsusenproductsaaccu Chek Inform IitoolkitOS0130802 Accu Chek Inform IIRose LeanoAinda não há avaliações
- Homoeopathic Drug Proving: Randomised Double Blind Placebo Controlled TrialDocumento9 páginasHomoeopathic Drug Proving: Randomised Double Blind Placebo Controlled TrialParag SharmaAinda não há avaliações
- Group 2 - Up-Fa1-Stem11-19 - Chapter IiDocumento7 páginasGroup 2 - Up-Fa1-Stem11-19 - Chapter IijamesrusselAinda não há avaliações
- Tle 7 - 8 - FisheryDocumento17 páginasTle 7 - 8 - FisheryRey JavierAinda não há avaliações
- Kinds of Blood. Differences Between Men and WomenDocumento11 páginasKinds of Blood. Differences Between Men and WomenTiagoSantosAinda não há avaliações
- 57 2 3 BiologyDocumento15 páginas57 2 3 BiologyAadya SinghAinda não há avaliações
- CP of Dexterous ConsultantsDocumento12 páginasCP of Dexterous ConsultantsDipankar GhoshAinda não há avaliações
- IELTS 1 Test IntroDocumento1 páginaIELTS 1 Test IntromichaelAinda não há avaliações
- 5 Problems (FSPR)Documento6 páginas5 Problems (FSPR)NMDNMSSDAinda não há avaliações
- Wellness at SeaDocumento9 páginasWellness at SeaRam Niwas ChauhanAinda não há avaliações
- Lecture 1 Introduction To Industrial HygieneDocumento40 páginasLecture 1 Introduction To Industrial Hygienesiti zubaidahAinda não há avaliações
- Ekolite KRYS 02 S PDS Rev 3Documento2 páginasEkolite KRYS 02 S PDS Rev 3Aldila Ratna OvrisadinitaAinda não há avaliações
- Attitude of EMPLOYEES in Terms of Compliance of Health and SafetyDocumento6 páginasAttitude of EMPLOYEES in Terms of Compliance of Health and SafetyJanice KimAinda não há avaliações
- Final Sheet MotalityDocumento69 páginasFinal Sheet MotalityAshima GabgotraAinda não há avaliações
- Hong Kong Dental Journal - Volume 2 - Number 2 - YuLeungDocumento1 páginaHong Kong Dental Journal - Volume 2 - Number 2 - YuLeungFebySiampaAinda não há avaliações
- Maslow TheoryDocumento10 páginasMaslow Theoryraza20100% (1)
- Gut: the new and revised Sunday Times bestsellerNo EverandGut: the new and revised Sunday Times bestsellerNota: 4 de 5 estrelas4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceNo EverandTales from Both Sides of the Brain: A Life in NeuroscienceNota: 3 de 5 estrelas3/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityNo EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityNota: 4 de 5 estrelas4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNo EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNota: 3.5 de 5 estrelas3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)No EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Nota: 4 de 5 estrelas4/5 (378)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeNo EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeAinda não há avaliações
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNo EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNota: 4.5 de 5 estrelas4.5/5 (15)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNo EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNota: 4.5 de 5 estrelas4.5/5 (516)