Você também pode gostar
- PANCREASDocumento27 páginasPANCREASJuan Jose Valladares SanchezAinda não há avaliações
- Tema: Biomoleculas Inorganicas - El Agua Docentes: Juan José Valladares Sánchez Hugo MurgaDocumento7 páginasTema: Biomoleculas Inorganicas - El Agua Docentes: Juan José Valladares Sánchez Hugo MurgaJuan Jose Valladares SanchezAinda não há avaliações
- Tema: Niveles de Organización de Los Seres Vivos Docentes: Juan José Valladares Sánchez Hugo MurgaDocumento9 páginasTema: Niveles de Organización de Los Seres Vivos Docentes: Juan José Valladares Sánchez Hugo MurgaJuan Jose Valladares SanchezAinda não há avaliações
- TAREA 5to Año S3Documento2 páginasTAREA 5to Año S3Juan Jose Valladares SanchezAinda não há avaliações
- Sílabo Inglés VII - MarzoDocumento5 páginasSílabo Inglés VII - MarzoJuan Jose Valladares SanchezAinda não há avaliações
- 22/10/19 TemaDocumento2 páginas22/10/19 TemaJuan Jose Valladares SanchezAinda não há avaliações
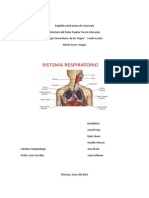
- Informe de Sistema RespiratorioDocumento10 páginasInforme de Sistema RespiratorioDailizMarinAinda não há avaliações
- Plan Vigilancia EntomológicaDocumento8 páginasPlan Vigilancia Entomológicacoco batanaAinda não há avaliações
- Protocolo EXPUESTO PERINATALDocumento4 páginasProtocolo EXPUESTO PERINATALstix6Ainda não há avaliações
- 2do Parcial MicrobiologiaDocumento2 páginas2do Parcial MicrobiologiaAlicia Mariaca ChipunaviAinda não há avaliações
- TP FilosofiaDocumento3 páginasTP Filosofiavaneclaudia100% (1)
- Inf Urinarias Vzla AdultoDocumento33 páginasInf Urinarias Vzla AdultoMacarenaAinda não há avaliações
- Matriz de RiesgosDocumento12 páginasMatriz de RiesgosWalter Orlando Lopez OlarteAinda não há avaliações
- 1363 Pares Biomagneticos PDFDocumento119 páginas1363 Pares Biomagneticos PDFJose Alexander Vasquez Leal100% (6)
- Abandono de La Terapia AntirretroviralDocumento17 páginasAbandono de La Terapia AntirretroviralDavid Martinez Barua100% (1)
- Antibiograma 2015 IIDocumento17 páginasAntibiograma 2015 IIErik Salvatierra LinaresAinda não há avaliações
- 93-Virus Respiratorio Sincitial BovinoDocumento2 páginas93-Virus Respiratorio Sincitial BovinoJanierth Guzman100% (1)
- Esquema de Inmunizacion 1Documento66 páginasEsquema de Inmunizacion 1Enfermeria UNPRGAinda não há avaliações
- Poliadenomatosis Pulmonar OvinaDocumento9 páginasPoliadenomatosis Pulmonar Ovinavoldur12Ainda não há avaliações
- Secuencias NuméricasDocumento12 páginasSecuencias NuméricasGiovanna Rodriguez100% (2)
- Triptico La ChancapiedraDocumento2 páginasTriptico La ChancapiedraSusana Urrutia100% (1)
- Equipos de Protección Básico y Especializado Según El Área de Trabajo y Circunstancias en Que Los UtilizanDocumento4 páginasEquipos de Protección Básico y Especializado Según El Área de Trabajo y Circunstancias en Que Los UtilizanLuz Karime Humanez Moreno100% (3)
- Alteraciones MicrobianasDocumento13 páginasAlteraciones MicrobianasElvia Gómez VillacortaAinda não há avaliações
- 697 20230418 20230418 AtenMedxServDocumento98 páginas697 20230418 20230418 AtenMedxServjoseAinda não há avaliações
- Trasfusión de Sangre y Sus Componentes Riesgos, Beneficios e IndicacionesDocumento23 páginasTrasfusión de Sangre y Sus Componentes Riesgos, Beneficios e IndicacionesMed_angelAinda não há avaliações
- Tarea 1 de Virus, Priones y ViroidesDocumento11 páginasTarea 1 de Virus, Priones y ViroidesAneudi Rafael Tineo AlmonteAinda não há avaliações
- Introducción LacteosDocumento2 páginasIntroducción LacteosDanielita Fernanda0% (1)
- Clase 1Documento49 páginasClase 1Fabián MoralesAinda não há avaliações
- Cuidados de EnfermeriaDocumento5 páginasCuidados de EnfermeriasalliqueAinda não há avaliações
- Contenido Programatico Infecciosas I 2020-2Documento14 páginasContenido Programatico Infecciosas I 2020-2Mike SanchezAinda não há avaliações
- Cuadro Sx. MononucleósicoDocumento3 páginasCuadro Sx. MononucleósicoOly VazquezAinda não há avaliações
- Mapa Conceptual Sobre La VirologíaDocumento1 páginaMapa Conceptual Sobre La VirologíaYamilaAinda não há avaliações
- StaphyloDocumento9 páginasStaphyloTosCof100% (3)
- Cuadro Comparativo OmDocumento1 páginaCuadro Comparativo OmSandra EducAinda não há avaliações
- Microbiologia de Los AlimentosDocumento18 páginasMicrobiologia de Los AlimentosArnold Gamarra Fuentes100% (1)
- NIFMuestra 1Documento13 páginasNIFMuestra 1Daniel Pereira100% (1)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadNo EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadNota: 4.5 de 5 estrelas4.5/5 (1833)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadNo Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadNota: 5 de 5 estrelas5/5 (199)
- Resetea tu mente. Descubre de lo que eres capazNo EverandResetea tu mente. Descubre de lo que eres capazNota: 5 de 5 estrelas5/5 (196)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualNo EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualNota: 4.5 de 5 estrelas4.5/5 (1700)
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaNo EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaNota: 5 de 5 estrelas5/5 (457)
- ¡Basta ya de ser un Tipo Lindo! (No More Mr. Nice Guy): Un Plan Probado para Que Obtengas Lo Que Quieras en La Vida El Sexo y El Amor (A Proven Plan for Getting What You Want in Love, Sex and Life)No Everand¡Basta ya de ser un Tipo Lindo! (No More Mr. Nice Guy): Un Plan Probado para Que Obtengas Lo Que Quieras en La Vida El Sexo y El Amor (A Proven Plan for Getting What You Want in Love, Sex and Life)Nota: 5 de 5 estrelas5/5 (47)
- Homo antecessor: El nacimiento de una especieNo EverandHomo antecessor: El nacimiento de una especieNota: 5 de 5 estrelas5/5 (1)
- Mujeres de luz: Guía para descubrir el poder de tu alma y su propósitoNo EverandMujeres de luz: Guía para descubrir el poder de tu alma y su propósitoNota: 4.5 de 5 estrelas4.5/5 (39)
- Joven a cualquier edad. El método definitivo para una vida larga, saludable y felizNo EverandJoven a cualquier edad. El método definitivo para una vida larga, saludable y felizAinda não há avaliações
- Las 6 necesidades de cada niño: Empoderar a padres e hijos a través de la ciencia de la conexiónNo EverandLas 6 necesidades de cada niño: Empoderar a padres e hijos a través de la ciencia de la conexiónNota: 4.5 de 5 estrelas4.5/5 (6)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceNo EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceNota: 5 de 5 estrelas5/5 (8)
- Escribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelNo EverandEscribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelNota: 5 de 5 estrelas5/5 (1)
- Fuerza de voluntad: Cómo practicar y dominar el autocontrolNo EverandFuerza de voluntad: Cómo practicar y dominar el autocontrolNota: 4.5 de 5 estrelas4.5/5 (91)
- Signos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososNo EverandSignos y símbolos: Desvelando el significado espiritual de los sellos angélicos, tótems y otros símbolos mágicos, sagrados y religiososNota: 5 de 5 estrelas5/5 (1)