Escolar Documentos
Profissional Documentos
Cultura Documentos
Anatomi Dan Fisiologi Pembuluh Darah PDF
Enviado por
Inyong BudionoTítulo original
Direitos autorais
Formatos disponíveis
Compartilhar este documento
Compartilhar ou incorporar documento
Você considera este documento útil?
Este conteúdo é inapropriado?
Denunciar este documentoDireitos autorais:
Formatos disponíveis
Anatomi Dan Fisiologi Pembuluh Darah PDF
Enviado por
Inyong BudionoDireitos autorais:
Formatos disponíveis
12
BAB II
TINJAUAN PUSTAKA
2.1 Anatomi dan Fisiologi Pembuluh Darah
2.1.1 Sistem Pembuluh Darah
Di antara berbagai organ tubuh, pembuluh darah mungkin merupakan salah satu
organ yang mempunyai peranan penting dan sistemnya sangat kompleks. Dikenal
dua sistem sirkulasi di mana pembuluh darah memegang peranan utama yaitu:
sistem sirkulasi sistemik dan sistem sirkulasi paru-paru (Guyton, 2000). Di setiap
sistem, masing-masing dikelompokkan menjadi 3 sistem yaitu sistem arterial,
sistem kapiler dan sistem venosa. Aorta adalah pembuluh darah besar bagian dari
sistem sirkulasi sistemik, yang keluar dari jantung dan berfungsi untuk membawa
darah jantung yang penuh berisi oksigen ke pembuluh arteri. Dari pembuluh aorta
yang besar kemudian bercabang menjadi beberapa pembuluh darah arteri yang
ukurannya lebih kecil dan membawa darah dari percabangan aorta keseluruh
tubuh, kecuali arteri paru-paru yang berfungsi sebaliknya (Guyton, 2000; High
beam encyclopedia, 2008; Farlex, 2008). Di target organ, pembuluh darah arteri
bercabang-cabang dan berakhir menjadi pembuluh darah yang lebih kecil yang
disebut dengan arteriol. Arteriol bekerja sebagai katup pengatur di mana darah
dilepaskan ke dalam kapiler. Kapiler adalah pembuluh darah terkecil yang
berfungsi untuk menukar cairan dan bahan gizi di antara darah dan ruang
interstisial (Guyton, 2000). Venula mengumpulkan darah dari kapiler-kapiler.
13
Secara berangsur-angsur mereka bergabung menjadi vena-vena yang makin lama
makin besar. Vena adalah pembuluh darah yang berfungsi sebagai penyalur yang
membawa darah dari jaringan kembali ke jantung (Guyton, 2000).
Secara histoanatomik, ketebalan dinding ketiga sistem ini berbeda, sesuai
dengan fungsi utamanya masing-masing. Aorta dan pembuluh darah arteri, karena
fungsinya untuk menyalurkan darah dari jantung ke seluruh tubuh, mengalami
tekanan yang tinggi. Sehingga pembuluh darah arteri memiliki dinding vaskuler
yang kuat dan darah mengalir dengan cepat ke jaringan-jaringan.
Arteriol yang berfungsi sebagai katup pengatur dari sistem arteri, memiliki
dinding otot yang kuat yang dapat menutup sama sekali arteriol tersebut sehingga
memungkinkannya untuk berdilatasi beberapa kali, dengan demikian dapat
mengubah aliran darah ke kapiler.
Kapiler, karena fungsinya sebagai penukar cairan dan bahan gizi, memiliki
dinding yang sangat tipis dan permeabel terhadap zat yang bermolekul kecil.
Selanjutnya dari kapiler darah kemudian berlanjut menuju venula-venula yang
kemudian bergabung menjadi pembuluh darah vena.
Vena, karena berfungsi mengalirkan darah kembali ke jantung, memiliki
tekanan dinding yang sangat rendah dan sebagai akibatnya dinding vena tipis.
Tetapi walaupun begitu, dinding vena berotot yang memungkinkannya untuk
mengecil dan membesar, sehingga vena mampu menyimpan darah dalam jumlah
kecil atau besar tergantung kepada kebutuhan badan.
14
Tabel 2.1 di bawah ini menunjukkan perbedaan ketebalan dinding
pembuluh darah, diameter lumen dan luas area sesuai dengan fungsinya dalam
sistem.
Tabel. 2.1. Tebal Dinding Pembuluh Darah, Diameter Lumen dan Luas
Penampang Lintang (Area) Pembuluh Darah
SISTEM
PEMBULUH
DARAH
TEBAL
DINDING
LUMINAL AREA
Aorta 2 mm 2.5 cm 4.5 cm
2
Arteri 1 mm 0.4 cm 20 cm
2
Arteriol 20 m 30 m 400 cm
2
Kapiler 1 m 5 m 4500 cm
2
Venol 2 m 20 m 4000 cm
2
Vena 0.5 mm 5 mm 40 cm
2
Vena Kava 1.5 mm 3 cm 18 cm
2
Sumber: Kardiologi Molekuler oleh Baraas F., 2006, hal. 187
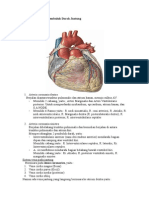
2.1.2 Struktur Dinding Pembuluh Darah
Dinding pembuluh darah terdiri dari 3 (tiga) lapisan, yaitu: lapisan terdalam yang
disebut sebagai tunika intima; yang ditengah disebut sebagai tunika media dan
yang terluar disebut sebagai tunika adventisia (Gambar 2.1). Tunika intima terdiri
dari selapis sel endotel yang bersentuhan langsung dengan darah yang mengalir
dalam lumen, dan selapis jaringan elastin yang berpori-pori yang disebut
membran basalis. Tunika media terdiri dari sel-sel otot polos, jaringan elastin,
proteoglikan, glikoprotein dan jaringan kolagen. Dalam keadaan biasa, jumlah
jaringan elastin yang membentuk tunika media aorta dan pembuluh darah besar
lainnya, lebih menonjol dibandingkan dengan otot polosnya. Sebaliknya di
15
pembuluh darah arteri lebih banyak dijumpai sel otot polos yang membentuk
tunika medianya. Perbedaan sel dalam tunika media menjadi tidak jelas (tidak bisa
dibedakan) bila sudah memasuki arteriol, bahkan tampaknya, dapat dikatakan
bahwa di dalam arteriol jaringan ikat dari tunika adventisia menjadi lebih dominan
(Guyton, 2000; Baraas F., 2006).
Gambar 2.1: Penampang Dinding Pembuluh Darah
Dalam dinding kapiler pembuluh darah, tidak didapatkan lagi lapisan
tunika media dan yang ada adalah lapisan sel endotel. Pada sistem venosa,
komponen tunika jumlahnya jauh lebih sedikit dibandingkan dengan sistem
arterial. Tunika media tidak begitu berkembang dan hanya terdapat pada vena
kava dan pembuluh darah vena besar lainnya. Pada vena-vena kecil dan venol,
hanya jaringan ikat tuna adventisia yang lebih dominan. Oleh karena itu sistem
venosa lebih mudah mengalami dilatasi yang ireguler dan menampung pembuluh
darah paling besar (Guyton, 2000; Baraas F., 2006).
Sumber: Hast, 2003
16
Elastin yang bersifat hydrofobik berperan dalam mempertahankan
elastisitas dinding pembuluh darah, sedangkan jaringan kolagen berperan dalam
mempertahankan struktur dan bentuk pembuluh darah. Jaringan kolagen pada
tunika media yang terdiri dari tiga tipe yaitu, tipe I dan tipe II mengandung sel-sel
fibril dengan diameter 20-90 nm, dan tipe III yang bersifat lebih elastik. Jaringan
ikat kolagen yang ada dalam tunika intima adalah jaringan kolagen tipe IV,
sedangkan yang tipe V ada di membran basal. Tunika adventisia yang merupakan
lapisan terluar bertindak sebagai pelindung dan terdiri dari banyak jaringan ikat,
saraf otonom, pembuluh darah limfe dan vasa vasorum (Guyton, 2000; Baraas F.,
2006).
2.1.3 Sel Endotel
Lapisan terdalam dari tunika intima, terdiri dari selapis sel yang disebut sel
endotel. Sel ini berbentuk pipih, poligonal dengan ukuran sekitar 10 x 50 m dan
tebalnya 1-3 m, dengan sumbu panjang sel sejajar dengan aliran darah (Baraas F.,
2006). Sel ini berada disemua struktur pembuluh darah mulai dari jantung sampai
dengan kepiler dan berhubungan langsung dengan aliran darah (Guyton, 2000,
Bruce A., et al., 2002, Baraas F., 2006).
Sel endotel berfungsi untuk mengatur aliran darah yang dipompa oleh
jantung menuju ke seluruh tubuh, begitu juga sebaliknya (Baraas F., 2006),
memiliki kemampuan yang luar biasa dalam mengadaptasikan dirinya, baik secara
jumlah maupun kemampuan mengatur untuk tujuan memenuhi kebutuhan lokal
(Bruce A, et al., 2002). Disamping itu sel ini, bilamana rusak akan mudah diganti
oleh adanya Vascular Endothelial Growth Factor (VEGF) (Mochida S etal.,
17
1998), hanya saja diperlukan waktu untuk proses regenerasi tersebut (Reidy etal.,
1986). Kelebihan inilah yang memberikannya kemampuan untuk memiliki fungsi
yang berbeda-beda sesuai dengan fungsi metabolik organ yang diembannya
masing-masing (Baraas F., 2006). Secara umum sel endotel memiliki 3 (tiga)
fungsi dasar, yaitu: Pertama, endotel berfungsi sebagai garis pertahanan utama
(barrier) terhadap hampir semua elemen asing yang mencoba invasi ke dalam
suatu organ; kedua endotel berfungsi sebagai tempat metabolisme dan
katabolisme senyawa-senyawa tertentu; dan ketiga, sel ini berfungsi sebagai
tempat sintesis berbagai senyawa vasoaktif yang diperlukan dalam
mempertahankan tonus pembuluh darah (Bruce, 2002, Bger, 2004), yaitu antara
lain sintesis berbagai mediator inflamasi, mediator proliferasi sel-sel subendotel
dan berbabagi faktor hemostasis lainnya (Guyton, 2000; Libby P., et al., 2002;
Najjar et al., 2005; Baraas F., 2006). Fungsi di atas disebabkan karena peran
utama sel endotel adalah mengendalikan sifat-sifat arteri seperti tonus vaskuler,
permeabilitas vaskuler, angiogenesis dan respon terhadap proses inflamasi (Najjar
et al., 2005)
Sel endotel mengeluarkan Oksida Nitrit (NO) yang berperan sangat
penting dalam mempertahankan tonus pembuluh darah khususnya untuk proses
relaksasi pembuluh darah (Libby P., et al., 2002; Bger, 2004; Najjar et al., 2005;
Baraas F., 2006). NO merupakan hasil dari proses perubahan L-Arginine menjadi
sitrulin yang dikatalisis oleh enzym Nitric Oxyde Syntase (NOS) yang termasuk
dalam kelompok sitokrom P-450. Telah dapat diidentifikasi 3 (tiga) isoform NOS
yaitu: neuralNOS (nNOS) yang berasal dari kromosom 7, inducible NOS (iNOS)
18
yang berasal dari kromosom 12 dan endothelial constitutive NOS (ecNOS) yang
berasal dari kromosom 17 (Baraas F., 2006).
Gaya gesek pulsatil (shear stress) darah dan dengan adanya ion Ca
2+
dari
luar sel dapat menyebabkan ecNOS menjadi aktif. Oleh karena itu produksi NO
oleh sel endotel distimulasi dan dipertahankan oleh faktor-faktor yang dapat
meningkatkan konsentrasi Ca
2+
intrasel, seperti gaya gesek pulsatil pada
permukaan sel-sel endotel yang selalu berlangsung disepanjang waktu, perubahan
keseimbangan berbagai molekul sinyal vasoaktif yang bersifat vasodilatatif
dengan molekul sinyal vasokonstruktif dan sebagainya (Baraas F., 2006).
Oksida Nitrit bekerja lokal (autokrin dan parakrin) oleh karena waktu
paruhnya sangat pendek dan segera bereaksi dengan air dan oksigen membentuk
nitrit dan nitrat. Oksida Nitrit ini selalu diproduksi dan didegradasi secara sangat
dinamik di dalam sel-sel endotel (Baraas F., 2006).
2.2 Penuaan Pembuluh Darah (Vascular Aging)
Dengan bertambahnya umur seseorang, proses menua yang terjadi sepanjang
hidup manusia akan tetap berlangsung. Seluruh organ beserta fungsinya, termasuk
pembuluh darah, juga mengalami proses menua. Penuaan organ ditandai dengan
berbagai perubahan struktur maupun fungsi. Berikut ini adalah pembahasan
tentang perubahan yang terjadi pada pembuluh darah sebagai akibat dari penuaan.
19
2.2.1 Definisi Penuaan yang Berkaitan dengan Pembuluh Darah
Umur merupakan faktor risiko dominan terhadap penyakit yang
menyerang pembuluh darah. Penuaan pembuluh darah dikaitkan dengan
perubahan struktur dan fungsi keberadaan pembuluh darah khususnya pembuluh
darah besar (Mengden, 2006; Nilson, 2008), seperti diameter lumen, ketebalan
dinding, peningkatan kekakuan dinding dan perubahan fungsi endotel (Mengden,
2006, Najjar et al., 2005). Pembuluh darah yang paling sering terkena adalah yang
bersifat elastis seperti aorta sentralis dan arteri carotis (Science Blog, 2003,
Lakatta, 2003; Najjar et al., 2005). Lumen pembuluh darah besar akan mengalami
dilatasi, dindingnya semakin tebal dan semakin kaku (Lakatta, 2003; Najjar et al.,
2005). Perubahan ini dipengaruhi oleh perubahan struktur, mekanika dan
biokimiawi dinding oleh karena faktor umur yang kemudian berakibat pada
menurunnya arterial compliance dan kakunya dinding (Jani & Rajkumar, 2006;
Laurent et al., 2006; Nilson, 2008). Najjar et al., 2005, yang mengutip pendapat
ORourke dan Nicholas, 2005, menyebutkan bahwa peningkatan kekakuan
dinding pembuluh darah adalah akibat dari siklus tekanan yang terus menerus dan
putaran yang berulang-ulang pada dinding elastis arteri, sehingga menekan
jaringan elastisnya untuk digantikan dengan jaringan kolagen. Selain itu Lakatta
dan Levy, 2003, dalam review artikelnya menyebutkan juga bahwa kekakuan
arteri ini berkaitan dengan pengaruh regulasi endotel terhadap tonus otot polos
arteri (Lakatta, 2003). Selanjutnya kemungkinan kekakuan dinding ini diperbesar
oleh adanya specific gene polymorphism (Hanon et al., 2001; Safar, 2005).
20
Perubahan struktur dan fungsi arteri yang berkaitan dengan umur pada orang sehat
dijelaskan dalam tabel di bawah ini.
Tabel. 2.2.: Perubahan Struktur dan Fungsi Arteri yang berkaitan dengan Umur
pada Manusia, Kera dan pada beberapa Mahluk Monogastrik
PARAMETER ARTERI
MENUA
HIPER-
TENSI
ATERO-
SKLEROSIS
MANUSIA
> 65 TH
KERA
15-20 TH
TIKUS
24-30 BL.
KELINCI
3-6 TH.
Lumen melebar + + + + ?
Kekakuan + + + + + ?
Collagen + + + + ?
Elastin + + + + ?
Disfungsi endotel + + + + + +
Intima menebal difus + + + + + +
Keterlibatan lemak - - - - +
jmlh VCMC + + + + + +
Macrophage + - - - + +
T sel + - - - + +
Matriks + + + + + +
Local Ang II-ACE + + + + + +
Disregulasi MMP + + + ? + +
MCP-1/CCR2 + + + + + +
ICAM ? ? + ? + +
TGFB ? + + ? + +
NADPH oxidase ? ? + ? + +
VEGF + ? ? + + +
NO bioavailability ? ? + + + +
panjang telomer + + + ? ? +
Hipertensi +
Aterosklerosis - - - +
Sumber: Najjar et al., 2005,
2.2.2 Perubahan Struktur Dinding
Di bawah mikroskop, menebalnya dinding arteri ditunjukkan oleh
ketebalan tunika intima dan media. Pada penelitian post mortem dijumpai bahwa
penebalan dinding aorta terjadi secara difus dan terutama terjadi di lapisan intima,
walaupun penelitian ini dilakukan di populasi dengan angka kejadian
aterosklerosis yang rendah (gambar 2.2) (Lakatta, 2003; Najjar et al., 2005).
21
Penebalan berjalan secara linier dan secara epidemiologis, dapat mencapai 2-3
kali lipat dari ukuran sebelumnya pada rentang usia 20-90 tahun walaupun tanpa
diikuti oleh plak aterosklerosis (Nagai et al., 1998, Lakatta, 2003; Najjar et al.,
2005). Penebalan terjadi secara tidak merata di dinding pembuluh darah, sangat
bervariasi, yang menunjukkan bahwa penebalan dinding oleh karena umur ini
sangat heterogen. Sehingga ada istilah mereka yang sukses atau yang
prosesnya dipercepat (Najjar, 2005).
Gambar 2.2: Penebalan dinding arteri yang menua. Sumber Najjar, 2005
Secara histologis, dinding intima yang menebal secara difus terdiri dari
matriks protein, collagen, glycosaminoglican dan sel otot polos vaskuler
(VSMCs). Otot polos vaskuler di tunika intima yang menua diduga berasal dari
22
tunika media yang kemudian bermigrasi ke intima, terjadi peningkatan ekspresi
dari molekul-molekul adesi yang di lapisan intima aorta dan peningkatan
adherence dari monocyt ke permukaan sel endotel (Najjar, 2005). Keberadaan
glycosaminoglican ini berperan penting terhadap regulasi beberapa sifat fungsi
arteri termasuk diantaranya permeabilitas vaskuler (Lakatta, 2003).
Pada sel endotel, perubahan yang terjadi seiring dengan meningkatnya
umur antara lain meningkatnya sel dengan inti polipoid, meningkatnya
permeabilitas endotel, perubahan susunan dan integritas cytoskeleton dari sel,
munculnya senescence-assosiated galactosidase staining dan ekspresi beberapa
penghambat siklus sel (Lakatta, 2003; Najjar, 2005).
Pada tunika media yang menua, perubahan yang menonjol antara lain
deposisi dari matriks protein ekstraseluler seperti Fibronectin dan type 2 matrix
metalloprotease (MMP-2) (Wang et al., 2003), yang mendorong degradasi
matriks protein dan memfasilitasi migrasi dari sel otot polos vaskuler (Pauly et al.,
1992).
Sel otot polos vaskuler di media aorta tikus yang tua ukurannya lebih besar
tetapi jumlahnya lebih sedikit bila dibandingkan dengan tikus dewasa (Najjar,
2005).
Karakteristik lainnya dari media yang menua adalah pergesaran isi dan
integritas dari struktur matriks protein yang disebut elastin dan collagen di mana
jaringan elastin akan berkurang dan hal ini berimplikasi pada semakin kakunya
dinding pembuluh darah oleh karena pertambahan umur (Najjar, 2005).
23
2.2.3 Perubahan Seluler Sel Arteri
Perubahan biomolekuler yang terjadi oleh karena penuaan pembuluh darah
bersumber terutama dari sel endotel dan selain itu juga dari tunika intima dan
media. Perubahan ini, pada manusia, mirip sekali dengan perubahan yang terjadi
oleh karena penyakit pembuluh darah seperti aterosklerosis ataupun hipertensi,
sehingga sering sekali perubahan dini dikatakan sebagai fase subklinik dari
terjadinya penyakit pembuluh darah tersebut (Lakatta, 2003; Najjar, 2005, Nilson,
2008).
2.2.3.1 Perubahan pada Sel Endotel (Struktur Dan Fungsi)
Sel endotel arteri yang tua mensekresi lebih banyak plasminogen activator
inhibitor-1 (PAI-1) yang mempermudah munculnya trombosis. Disamping itu,
pada arteri yang menua ini terjadi peningkatan dari endotelin dan
vasoconstructing growth factors yang diproduksi oleh sel endotel, antara lain
Angiotensin II. Sebaliknya faktor vasodilatasi, seperti NO, prostacyclin dan
endothelium hyperpolarising factor menurun. Perubahan yang terjadi di pembuluh
darah karena penuaan ini memberikan suasana aktif baik secara enzimatis maupun
metabolik terhadap terjadinya penyakit pembuluh darah seperti aterosklerosis
(Najjar, 2005)
Panjang telomer dalam endotel merupakan marker untuk penggantian sel
dan ini berbanding terbalik dengan umur, gradasi aterosklerosis dan tekanan nadi
(Lakatta, 2003). Hilangnya telomer pada arteri yang menua menginduksi disfungsi
endotel di sel endotel vaskuler, sedangkan inhibisi terhadap pemendekan telomer
24
akan menekan proses disfungsi sel yang berkaitan dengan umur (Minamino et al.,
2002).
Endotel disini menunjukkan telomer yang lebih pendek dan aktivitas
telomerase reverse transcriptase juga tertekan menyebabkan proses glikasi dari
jaringan colagen yang dapat menginduksi munculnya perubahan fenotip sel
endotel seperti sel senescence. Advanced glycation end products yang
terakumulasi oleh karena umur akan mengaktivasi NAD(P)H oksidase yang akan
meningkatkan produksi anion superoksida. Menyatunya Advanced glycation end
products (AGE) dan reseptornya (RAGE) di sel endotel akan memicu pengerahan
dan aktifasi sel inflamasi yang meningkatkan trombogenesis dengan cara
menstimulir agregasi platelet (Wautier, 2004). Oleh karena itu proses menua dapat
dikatakan sebagai proses inflamasi kronis yang lamban dan ditandai oleh
munculnya sitokins pro inflamasi, seperti TNF-, IL-6 and NFB (Donato et.al.,
2009)
2.2.3.2 Perubahan pada Tunika Intima
Pada tunika intima dinding arteri binatang primata dan binatang pengerat
yang tua didapatkan penebalan yang difus, walaupun kedua binatang ini tidak
menderita aterosklerosis (Li Z. et al., 1999; Asai et al., 2000).
Diantara lapisan intima yang menebal ini tingkatan dari inflamatory
chemokine yaitu monocyte chemocontractant protein-1 (MCP-1) dan reseptornya
juga meningkat yang menyebabkan penebalan dan invasi sel otot polos (VSMCs)
ke dalam intima (Spinetti G. et al., 2004). Najjar et al. (2005) yang mengutip
25
laporan dari Boring et al., (1998) menjelaskan bahwa hal ini berimplikasi terhadap
patogenesis dari proses aterosklerosis (Boring etal, 1988; Najjar et al., 2005).
Molekul inflamasi ini (termasuk didalamnya MCP-1) diproduksi dan disekresi
oleh sel endotel dan sel otot polos (VSMCs), sedangkan pada binatang tidak
dijumpai sel inflamasi tradisional seperti lekosit di dinding pembuluh darah aorta
(Najjar, 2005).
Disamping itu, ekspresi dan aktivitas faktor pertumbuhan multifungsi
(multifunctional growth factor), transforming growth factor-
1
(TGF-
1
) di intima
juga meningkat (Wang & Lakatta, 2002). Faktor pertumbuhan inilah yang
mengatur replikasi dan sintesis komponen matriks ekstra sel dan memberikan
respon terhadap injury (Roberts and Sporn ,1990, Shah M etal., 1995).
Selanjutnya, keberadaan Nitric Oxide (NO) di intima arteri yang menua
juga berkurang, sedangkan aktivitas NADP(H) oksidase dan produksi reactive
oxygen species juga meningkat (Cernadas et al., 1998; Hamilton et al., 2001).
Keadaan ini akan memberi kesempatan terjadinya peroksidasi lipid dan modifikasi
protein karena oksidasi juga.
2.2.3.3 Perubahan di Tunika Media
Pada tunika media yang menua, perubahan yang menonjol antara lain
deposisi dari matriks protein ekstraseluler seperti Fibronectin dan type 2 matrix
metalloprotease (MMP-2) (Wang et al., 2003) yang mendorong degradasi matriks
protein dan memfasilitasi migrasi dari sel otot polos vaskuler (Pauly et al., 1992).
26
Sel otot polos di tunika media tampaknya mengalami proses modulasi
penotip ke arah dedifferentiated dan synthetic state. Menurut Najjar, 2005, proses
migrasi sel otot polos dari media ke intima menjadi lebih masuk akal yaitu oleh
karena peningkatan jumlah sel otot polos di dinding intima arteri sentral yang
menebal seiring dengan pertambahan usia. Selanjutnya, bila terjadi arterial injury
pada tikus yang tua, sel otot polos disini akan bertumbuh menyertai formasi
neointimal yang berlebihan dan aselererasi respon remodeling dinding pembuluh
arteri. Pertumbuhan yang berlebihan ini melambangkan faktor intrinsik pada
dinding pembuluh darah (Hariri et al., 1988)
Penurunan jaringan elastin oleh karena umur lebih disebabkan karena
menurunnya sintesis elastin. Penurunan ini disebabkan oleh karena represi
terhadap ekspresi gen elastin oleh B-Myb dan degradasi serabut elastin yang
kejadiannya dipercepat oleh adanya proses enzymatis seperti MMP-2 yang
tingkat dan aktivitasnya meningkat dengan bertambahnya usia (Wang et al., 2003).
Fragmen elastin yang dihasilkan akan berinteraksi dengan reseptor elastin-laminin
yang ada di beberapa permukaan sel termasuk sel endotel dan sel otot polos dan
menginduksi motilitas dan proliferasinya.
Sebaliknya pada binatang ini terjadi penimbunan kolagen tipe I dan III di
tunika media pembuluh darah yang sudah menua (Wang et al., 2002). Jaringan
collagen yang berdekatan akan mengalami glikasi enzimatis dan oksidasi sehinga
menghasilkan glycation end products (Vaitkevicius et al., 2001). Yang membuat
dinding arteri menjadi kaku.
27
2.3 Teori Penuaan Pembuluh Darah
Patofisiologi penuaan pembuluh darah dijelaskan oleh beberapa teori yang
berjalan seiring dan saling mempengaruhi. Teori-teori yang dapat menjelaskan hal
tersebut antara lain
2.3.1 Teori Stress Oksidatif
Sel endotel pembuluh darah mempunyai peranan paling penting dalam
merespon setiap invasi molekul antigen, sehingga dapat dikatakan bahwa sel
endotelah yang menjaga dan memelihara keseimbangan tubuh manusia. Setiap
faktor yang menyebabkan perubahan pada permukaan membran sel, secara
otomatis akan direspon oleh sel endotel dalam upaya untuk mengembalikan atau
mempertahankan keseimbangan itu kembali (Baraas F., 2006). Untuk
mengantisipasi keadaan ini, sel endotel senantiasa memproduksi faktor relaksasi
dan penghambat pertumbuhan seperti Nitric Oxide (NO), prostacyclin,
endothelium-derived hyperpolarizing factors (EDHF) dan faktor vasokonstriksi
dan faktor promosi pertumbuhan seperti superoxide anions (O
2
-
), endoperoxides,
thromboxane A
2
, endothelin (ET)-1, Angiotensin II, secara seimbang (Vanhoutte,
2002).
Invasi molekul yang mengenai endotel ini disebut sebagai stres oksidasi,
di mana stres ini dapat menimbulkan kerusakan pada sel endotel ini. Stress ini
dapat disebabkan oleh perubahan tekanan gaya gesek pulsatil pada permukaan sel
endotel, iritasi bahan kimiawi, trauma fisik, infeksi, polusi, asap rokok, hipoksia,
fenomena iskemia-reperfusi dan fenomena dismetabolik (Baraas F., 2006).
28
Setiap saat sel endotel akan selalu mengalami proses ini sehingga dapat
dikatakan peristiwa ini merupakan salah satu mekanisme dasar dari terjadinya
kerusakan endotel. Stress oksidasi yang kronis walaupun ringan akan memotong
integritas telomere yang menyebabkan sel endotel mengalami senescence secara
prematur (Kurz et al., 2004).
Dengan bertambahnya umur, keberadaan faktor pembuluh darah
yang baik (relaksasi dan penghambat pertumbuhan) akan menurun
dibandingkan dengan faktor jelek (kontraksi dan promotor pertumbuhan) yang
meningkat (Brandes et al., 2005). Produksi O
2
-
yang meningkat tampaknya akan
mengikis reaktivitas dari endothelium dependent nitrovasodilator di dinding aorta
dan berakibat berkurangnya daya untuk relaksasi (Brandes et al., 2005).
Keberadaan Oksida Nitrit (NO) pada penuaan arteri menurun tetapi
aktivitas eNOS masih kontroversi. Beberapa studi menyebutkan bahwa ekspresi
Endothelium Nitric Oxide Synthase (eNOs) dan NO menurun sehingga
mengakibatkan terjadinya stress oksidasi (Csiszar et al., 2002), tetapi beberapa
studi menyebutkan ekspresi eNOS meningkat dan peningkatan ini dirangsang oleh
tingginya O
2
-
(van der Loo et al., 2000). Meningkatnya O
2
-
besar kemungkinan
oleh karena rangsangan NaD(P)H oksidase yang memang meningkat pada
pembuluh darah yang menua (Csiszar et al., 2002). O
2
-
merangsang eNOS untuk
memproduksi NO , tetapi tingginya rangsangan untuk memproduksi NO dan oleh
O
2
-
yang terus menerus akan membentuk peroxynitrite (ONOO
-
) dan uncoupling
dari Enos (gambar 2.3). Peroxynitrite inilah yang mengikis ketersediaan NO dan
menghambat kerja antioksidan Manganese Superoksida Dismutase (MnSOD) di
29
dalam plasma yang sedianya akan mengubah Radikal Superoksida (O
2
-
) segera
menjadi H
2
O
2
(van der Loo et al., 2000, Griendling et al., 2003, Brandes et al.
2005, Schiffrin, 2008), untuk selanjutnya oleh enzym Catalse dan GSH H
2
O
2
akan
segera dirubah menjadi H
2
0 dan O
2
(Griendling and FitzGerald, 2003). Disamping
itu Peroxynitrit (ONOO
-
) juga merupakan mediator penting dari peroksidasi
lemak termasuk oksidasi LDL dan proses nitrasi protein yng berujung pada
aterogenesis (Griendling et al., 2003).
Peningkatan umur berlanjut dengan disfungsi mitokondria yang
disebabkan oleh karena kerusakan DNA didalamnya. Sepanjang hidup manusia,
mitokondria selalu memproduksi Superoxide anion (O
2
-
) dengan jumlah yang
berbeda-beda sesuai dengan lokasinya dan DNA dalam mitokondria selalu
terpapar terhadap stres oksidatif yang berakibat pada kerusakan DNA. Biasanya
O
2
-
cepat diubah menjadi H
2
O
2
oleh manganese superoxide dismutase (MnSOD),
untuk selanjutnya diubah menjadi H
2
O dan O
2
tadi. Sebagai akibat dari produksi
O
2
-
yang terus menerus, maka jumlah mitokondria akan berkurang, ekspresi
proteinnya akan melemah seperti format disfungsi protein yang mengarah pada
deplesi energi seluler yang berlanjut dengan terbentuknya radikal lanjutan seperti
peroxynitrit (ONOO
.-
). Dalam pembuluh darah, peroxynitrit yang menonaktifkan
MnSOD di Mitokondria akan merangsang terbentuknya kembali O
2
-
dalam
mitokondria (Griendling et al., 2003, Brandes et al., 2005).
Kerusakan DNA tidak terbatas di mitokondria saja dan ada satu enzym
yang bekerja dalam melabel DNA yang rusak yaitu Poly(ADP-ribose)polymerases
30
(PARP) yang ternyata juga terlibat dalam penipisan endotel yang menyebabkan
relaksasi pada penuaan
Beberapa enzym yang ikut dalam proses pembentukan radikal antara lain,
NO sintase, xantin oksidase, dan walaupun kejadiannya kurang, NADPH oxidase
juga dapat dikatakan sebagai sumber dari pembentukan radikal karena enzym ini
dapat mengaktifkan GTPase NADPH oxidase subunit Nox4 yang ternyata mampu
membentuk O
2
-
dan sel senescence (Brandes et al., 2005). Bahkan NADPH
oxidase dianggap sebagai sumber terpenting dari terbentuknya O
2
-
dalam dinding
pembuluh darah (Schiffrin, 2008). Di pihak lain, NADPH dibentuk oleh
vasoactive hormones dan protein G-protein rac-1 yang berat molekulnya kecil
(Griendling et al., 2003). Meningkatnya peroksida ini dapat menjadi persisten
Gambar 2.3:
Proses munculnya
radikal bebas dalam
endotel yang menua
Sumber: Brandes et al., 2005
31
oleh karena pemberian kholesterol yang tinggi secara terus menerus, dan hal ini
berakibat pada tertekannya aktivitas NO (Griendling and Fitzgerald, 2003).
Disamping kerusakan atau modifikasi DNA, pembentukan radikal bebas
ini juga akan memodifikasi lemak dan protein, meregulasi beberapa bagian dari
gen seperti adhesion molecules, chemotactic factors, enzim antioksidan,
vasoactive substances. Meningkatnya ROS berakibat pada oksidasi LDL, bila
konsentrasi lemak dalam darah tetap tinggi (Berliner etal, 1985, Griendling, 2003),
degradasi kolagen yang disebabkan oleh termodifikasinya aktivitas Matrix
Metalloproteinase (MMPs) seperti MMP-2 dan MMP-9 (Griendling et al., 2003).
2.3.2 Teori Inflamasi (Respon Imunologi)
Di dalam tunika intima yang menebal pada tikus yang tua ditemukan
beberapa sitokin seperti Transforming Growth Faktor-beta (TGF-), Interstitial
Cell Adhesion Molecule (ICAM-1), dan enzym zinc-dependent endopeptidase
type-2 metalloproteinase (MMP-2) (Lakatta, 2003). Hal ini menunjukkan adanya
pengaruh proses inflamasi pada penuaan pembuluh darah.
Proses inflamasi ini merupakan respon terhadap lesi yang terjadi pada sel
endotel (vascular respon to injury) oleh karena tekanan dan oleh karena gaya
gesek dari darah. Ada dua jalur respon terhadap lesi endotel yaitu melewati jalur
NO yang sudah dibicarakan sebelumnya dan jalur imunologik. Respon
imunologik ini diawali dengan peningkatan sekresi sitokin, sekresi faktor-faktor
koagulasi dan reaktivasi platelet, yang sangat besar peranannya dalam proses
trombosis akut (Baraas F., 2006).
32
Respon imunologik yang akut diawali dengan keluarnya sitokin
proimunologik melalui aktivasi faktor transkripsi NFB (Nuclear Faktor Kappa-
B). Faktor ini kemudian bergerak ke dalam inti sel untuk berikatan dengan DNA
untuk ekspresi beberapa gen target yang mengkode sitokin-sitokin, protein fase
akut dan sebagainya. Beberapa sitokin yang diekspresi antara lain TNF-, IL1-
17, IFN-. (Baraas F., 2006). Sitokin-sitokin inilah, pada sel otot polos pembuluh
darah yang menebal, merangsang disekresinya MMP-2 dan terjadi proliferasi
Platelet Derive Growth Factor (PDGF) dan peningkatan TGF- yang merangsang
formasi matriks fibronectin dan collagen (Lakatta, 2003). Respon lokal yang
muncul akibat ekspresi sitokin tersebut antara lain munculnya ICAM-1, VCAM-1
(Vascular Cell Adhesion molecule-1) dan PECAM-1 (Platelet Endothelial Cell
Adhesion Molecule-1) pada dinding pembuluh darah (Libby et al., 2002, Baraas F.,
2006).
Bilamana proses berlangsung kronis, berulang-ulang dan berkepanjangan,
maka respon proinflamasi dan protrombosis yang semula bersifat akut akan
berubah menjadi kronis. Akan terjadi infiltrasi sel-sel leukosit, terutama sel
monosit, ke bawah jaringan subendotel dan kemudian berubah menjadi sel
makrofag. Sel ini kemudian akan menelan berbagai sisa LDL yang teroksidasi
membentuk sel busa dan kemudian menjadi ateroma (Baraas F., 2006).
Nuclear faktor (NF)- B (respon elemen interferon- ), dan motif ikatan
DNA NFinterleukin-6 (IL-6) berada pada promoter dari Receptor Glication End
Product (RAGE).
NF- B mengatur ekspresi seluler dari RAGE, menghubungkan
RAGE dengan respon inflamasi (Li & Schmidt, 1997). RAGE adalah receptor dari
33
Advanced Glycation End Product (AGE) yang merupakan modifikasi protein atau
lipid yang mengalami glikasi nonenzymatik dan teroksidasi setelah kontak dengan
gula aldosa (Singh et al., 2001). AGE dibentuk oleh 3 faktor kunci yaitu
kecepatan protein turnover untuk proses glycoxidation, tingginya kadar glukosa
darah,
dan keadaan lingkungan yang menimbulkan stress oksidasi. AGE dapat
mengubah sifat-sifat dari matrix
protein collagen, vibronectin, dan laminin,
melalui ikatan AGE-AGE
intermolecular covalent, atau cross-linking. Cross-
linking AGE pada collagen tipe I dan elastin menyebabkan peningkatan matrik
ekstra seluler yang berdampak pada meningkatanya kekakuan pada vasculature
(Goldin et al., 2006).
AGE yang beredar di sirkulasi dapat berinteraksi dengan RAGE di endotel
(gambar 2.4.) yang menimbulkan pertuberasi sifat-sifat sel seperti peningkatan
regulasi transkripsi faktor NF- B dan kemudian dapat mentranskripsi beberapa
target gen seperti endothelin-1,
vascular cell adhesion molecule-1 (VCAM-1),
intercellular adhesion
molecule-1 (ICAM-1), E-selectin, tissue faktor,
thrombomodulin,
vascular endothelial growth faktor (VEGF), dan sangat mungkin
proinflammatory
sitokins, seperti IL-1 , IL-6, and tumor necrosis faktor-
(Yamagishi etal., 1998; Wautier & Guillausseau, 2001; Basta et al. 2004) .
Disamping itu NF- B adalah essensial dalam menginduksi adhesi monosit dan
migrasi oleh AGE (Morigi et al., 1998) dan meginduksi hiperpermeabilitas sel
endotel (Basta et al., 2004).
34
Sumber: Wautier & Guillausseau, 2001, Diabetes Metab, 27,
Gambar 2.4: Proses Glikasi yang menimbulkan disfungsi endotel
AGE yang ada di basal membran menghambat migrasi monosit dan menginduksi
proses yang disebut sebagai apostaxis. AGE juga berkontribusi terhadap ekspresi
reseptor oxidized LDL (OxLDL) pada monosit yang menjadi makrofag yaitu
macrophage scavenger receptor class A and CD36 sehingga dengan demikian
meningkatkan uptake OxLDL yang menimbulkan terbentuknya Sel Busa (Foam
Cell) (Iwashima et al., 2000) .
2.3.3 Kerusakan Endotel
Sel endotel pembuluh darah yang sehat berfungsi untuk mengatur
homeostasis dari sistim vaskular. Disfungsi endotel dianggap sebagai awal yang
sangat penting bagi munculnya kerusakan aterotrombotik yang mengarah kepada
munculnya penyakit pembuluh darah (Baraas F., 2006).
Sel endotel yang sehat mensekresi substrat yang bersifat proteksi dan yang
terpenting disini adalah Nitric Oxide (NO). Dalam keadaan sakit atau injury
35
seperti munculnya faktor risiko kardiovaskuler, aktivasi endotel mengarah kepada
disekresinya faktor kontraksi yang bekerja justru merugikan dan bahkan mengikis
NO. Disamping itu faktor kontraksi ini juga mencegah dan mengaktivasi berbagai
mekanisme yang berkaitan dengan aterotrombosis seperti yang telah dibahas
sebelumnya (Versari et al., 2008)
Begitu juga dalam kondisi semakin tua, fungsi sel endotel dalam
mempertahankan homeostasis juga semakin berkurang, yang disebabkan terutama
oleh karena adanya stress oksidatif oleh Reactive Oxygen Species (ROS). Stres
oksidatif ini menyebabkan dipercepatnya pemendekan telomere yang dinyatakan
melalui analisa terhadap terminal restriction fragment (TRF) dan hilangnya
integritas telomere inilah yang yang memicu proses senescence prematur dari sel
endotel (Kurz et al., 2004)
2.3.4 Teori Pemendekan Telomere
Pada endotel arteri yang tua didapatkan telomere yang mengalami keausan
dan supresi terhadap aktivitas telomerase (Chang & Harley, 1995, Lakatta, 2005).
Panjang telomere berhubungan dengan penggantian sel invivo maupun invitro,
sehingga dengan peningkatan keausan telomere diikuti dengan penurunan proses
penggantian sel dan diikuti dengan perubahan ekspresi gen yang tampak dalam sel
yang mengalami senescence (Chang & Harley, 1995, Najjar et al., 2003, Nilson,
2008) Proses ini terutama terjadi di pembuluh arteri yang mengalami tekanan
hemodinamika yang lebih banyak (Chang & Harley, 1995). Bila senescence cell
dari endotel ini terakumulasi di daerah yang proses penggantian selnya tinggi,
36
menurunnya kemampuan memecah diri akan membuat tunika media menjadi satu
lapis saja dan mengeksposnya terhadap faktor mutasi mitogenic dan adhesi yang
berkontribusi terhadap munculnya plak aterogenik (Chang & Harley, 1995,
Minamino et al., 2002).
Kurz dan kawan-kawan (2004) menyatakan bahwa pemendekan telomere,
disebabkan oleh stres oksidasi yang ringan tetapi berulang-ulang secara kronis.
Dijelaskan di sini bahwa di dalam sel endotel yang normal, stres oksidasi yang
ringan tetapi kronis yang diinduksi oleh pengaruh pertahanan antioksidan seperti
glutation, justru mengaselerasi erosi telomere dan onset sel yang mengalami
senescence.
Di samping teori yang disampaikan oleh Chang dan Harley, 1995, Wautier
(2004) juga mengemukakan bahwa pada telomere yang memendek dan aktivitas
telomerase reverse transcriptase yang tertekan akan menyebabkan proses glikasi
dari jaringan kolagen yang dapat menginduksi munculnya perubahan penotip sel
endotel seperti sel senescence. Advanced glycation end products (AGE) yang
terakumulasi oleh karena umur akan mengaktivasi NAD(P)H/oksidase yang akan
meningkatkan produksi anion superoksida. Menyatunya Advanced glycation end
products dan reseptornya di sel endotel akan memicu pengerahan dan aktifasi sel
inflamasi yang meningkatkan thrombogenesis dengan cara menstimulir agregasi
platelet (Wautier & Schmidt, 2004).
2.4 Penuaan Pembuluh Darah dan Aterosklerosis
Penuaan pembuluh darah berkaitan sangat erat dengan ateroskelrosis
bahkan dapat disebutkan bahwa faktor risiko utama dari aterosklerosis adalah
37
pembuluh darah yang menua, atau dengan kata lain aterosklerosis pada usia muda
adalah penuuan dini pembuluh darah.
Faktor risiko dan perjalanan dari proses penuaan pembuluh darah tidak
berbeda dengan proses terjadinya aterosklerosis. Yang berbeda hanya waktunya di
mana umur merupakan faktor utama yang independen terhadap lamanya proses
interaksi, sedang faktor lainnya dapat mempercepat atau megurangi kecepatan
proses interaksi tersebut. Secara diagramatis proses dapat dijelaskan dalam
gambar 2.5.
Gambar 2.5: Proses penuaan pembuluh darah yang ditandai dengan kerusakan sel
endotel berakhir pada aterosklerosis
(Modifikasi dari: Baraas F., 2006, Kardiologi Molekuler, Yayasan
Kardia Iqratama, hal 200)
Gaya gesek
pulsatil
Iskemia
Reperfusi
Infeksi
Polusi Merokok
Alograf
Bahan kimia
Kelebihan makan
Obesitas
Diabetes
Dislipidemia
Hipertensi
Stress
Oksidatif
ROS
Peroksidasi lipid
pada membrane sel
Endotel, LDL dll
Luka endotel,
LDL teroksidasi
Disfungsi endotel
Respon NO
Respon imunologik
Trombosis
Aterosklerosis
Bahan Fisika
38
2.4.1 Stress oksidasi dan aterosklerosis
Seperti disebutkan diatas, sel endotel pembuluh darah selalu mengalami
stress oksidatif yang sebetulnya merupakan suatu proses yang alami. Stres ini
dapat meningkat oleh beberapa keadan antara lain: Gaya gesek pulsatil pembuluh
darah, Iskemia reperfusi, infeksi, polusi khususnya oleh asap rokok, Alograf,
Bahan-bahan kimiawi, bahan-bahan fisika, hipertensi, dislipidaemia, obesitas,
makan berlebihan (Baraas F., 2006). Plasma kolesterol, khususnya kolesterol yang
teroksidasi disebutkan berkontribusi menghasilkan radikal bebas di sel endotel
dinding pembuluh darah. LDL yang teroksidasi menjadi OxLDL menstimuli
terbentuknya O
2
-
yang menimbulkan apoptosis pada dinding sel vaskuler (Awal
Prasetyo dan Udadi Sadhana, 2006; Bahorun etal., 2006).
Gambar 2.6: Mekanisme terbentuknya Foam Cell dalam proses Aterosklerosis
Modifikasi dari Awal Prasetyo dan Udadhi Sadhana, 2006
ROS
39
Stress oksidatif (ROS) (gambar 2.6) dapat menimbulkan peroksidasi
lemak pada dinding sel termasuk sel endotel yang menimbulkan jejas pada sel
tersebut (Endothelial injury). Jejas di dalam sel endotel inilah yang ditunjukkan
oleh munculnya respon NO dalam bentuk peningkatan yang diikuti dengan respon
imunologik. Di samping itu ROS yang berlangsung terus menerus walaupun
ringan dapat mengaselerasi sel senescence. Kejadian ini bila berulang terus akan
memendekkan telomere sel endotel disamping juga memicu munculnya
Advanced glycation end products (AGE), dan AGE yang terakumulasi oleh
karena umur akan mengaktivasi NAD(P)H oksidase yang akan meningkatkan
produksi anion superoksida disamping juga dapat mengaktifkan monosit menjadi
macrophage. Proses berlangsung terus yang menimbulkan disfungsi endotel yang
mengarah kepada trombosis (Baraas F., 2006). NAD(P)H oksidase tidak hanya
meningkat oleh karena meningkatnya umur, tetapi terjadi juga pada keadaan
patofisiologis yang menyebabkan penyakit kardiovaskuler seperti
hipercholesterolemia, hipertensi dan diabetes (Csizar et al., 2002). Macrophage
dan lemak terutama low-density lipoprotein (LDL) terakumulasi di daerah injury
di mana LDL akan teroksidasi dan dimakan oleh macrophage dan atau
macrophage sendiri juga teroksidasi sehingga memproduksi foam cell, yang dapat
berkembang menjadi plak aterosklerosis. Berliner et al., 1995 menyebutkan
bahwa proses oksidasi lemak lebih disebabkan oleh lamanya LDL yang
terakumulasi di endotel dibandingkan dengan tingginya kadar LDL dan LDL yang
teroksidasi akan bersifat toksik buat macrophage yang dapat berkontribusi
40
terhadap memburuknya proses inflamasi dan formasi pusat nekrosis pada stadium
yang lanjut.
Disamping itu macrofag foam cell yang terbentuk oleh karena asupan
lemak yang tinggi tidak hanya berasal dari teroksidasnya LDL menjadi oxLDL
tetapi juga dapat berasal dari penyerapan Very Low Density Lipoprotein (VLDL)
dan Agregated LDL (AgLDL) (Persson etal., 2006).
2.4.2 F2-Isoprostane untuk mengukur Stress Oksidasi dan Disfungsi
Endotel
Dalam keadaan normal lemak akan megalami oksidasi menjadi radikal
yaitu lipid peroksida molekul lemak radikal (
.
L) dan atau peroksida lemak (LOO
.
).
Kedua molekul ini bersifat radikal (Best, 2003)
.
OH + LH =>
.
L + H
2
O
.
L + O
2
=> LOO
.
LOO
.
+ LH => LOOH +
.
L
Fe
++
+ LOOH + H
+
=> Fe
+++
+
.
OL + H
2
O
Proses terjadi di dalam mitochondria dimana kedua molekul radikal tersebut
bersifat sangat reaktif dan dapat menembus hinga ke sitoplasma.
Oksidasi lemak dapat diukur dari metabolit yang dihasilkannya.
Peroksidasi lemak menghasilkan metabolit metabolit teroksidasi seperti keto,
hidroksi, epoksi dan beberapa asam lemak lebih pendek lainnya (Velasco et al.,
2004). Hydroperoksida yang dihasilkan oleh Asam Arachidonat kemudian dapat
dideteksi sebagai F2-isoprostan. Isoprostan adalah produk yang dihasilkan dari
41
jalur cyclooxygenase oleh mekanisme yang di katalisis oleh radikal bebas. Jadi,
metabolit ini dihasilkan sebagai ester phospolipid dari proses penyusunan kembali
stereo dan struktur isomer derivat PGH2 yang diproduksi oleh proses
cyclooxigenase asam arachidonat yang diinduksi oleh radikal bebas (Morrow J.D.
et al. 1990). Secara invitro F2-isoprostan terbentuk dari peroksidasi lemak LDL
oleh Cu2+ atau juga oleh pengoksida larut dalam air lainnya seperti 2,2-azo-bis-2-
amidinopropane (AAPH)(Lynch etal., 1994).
Asam Arachidonat yang termasuk dalam seri (n-6) asam lemak tak jenuh
jamak (PUFA), secara enzymatis melalui siklus Cyclooxygenase (COX) dirubah
menjadi bioaktif Prostaglandin (PGF
2
, PGE
2
, PGl
2
dan lain-lain) dan
Thromboxane (TX). Tetapi secara non enzymatik melalui jalur radikal bebas
dapat juga dirubah menjadi isoprostan (8-Iso-prostaglandin F
2
(8-iso-PGF
2
; F
2
-
isoprostanes), dan diketahui senyawa ini merupakan biomarker yang reliable
untuk mengukur stress oksidasi baik di eksperimen maupun dalam study klinik
(Nlsn C etal., 2006).
Senyawa ini sudah dibuktikan dapat menjadi indeks yang sangat berarti
untuk melihat peroksidasi lemak secara invivo (Morrow J.D. etal., 1998). Salah
satunya dipakai untuk mengukur munculnya radikal bebas oleh karena oksidasi
Low Density Lippoprotein (oxLDL) di dalam plasma darah (Dhawan etal., 2004),
menggambarkan proses kalsifikasi pada arteri koroner (Gross M. etal., 2005) dan
Lavi dalam artikelnya mengkaitkan antara F2-isoprostan, stress oksidasi dan
kerusakan endotel dimana disebutkan bahwa F2isoprostan menggambarkan
kerusakan endotel yang disebabkan oleh radikal bebas endogen (Lavi S etal.,
42
2008). Sehingga saat ini F2-isoprostan digunakan sebagai marker yang spesifik
dan ajeg dalam menggambarkan stress oksidasi secara invivo (Milne etal., 2005;
Comporti M. et al., 2008).
Robert Arjuna dalam disertasinya pada tahun 2007, menyebutkan bahwa
F2-isoprostan meningkat pada serum tikus yang menderita dislipidaemia menjadi
(28.06 8.26 ng/dl) dibandingkan tikus yang normal (21.99 5.70 ng/dl).
Akan tetapi ada kelemahan dalam menggunakan F2-isoprostan sebagai
marker invivo adalah kecepatan eliminasinya. Haliwell B. dan Lee C.Y.J (2010)
dalam reviewnya menyebutkan bahwa F2isoprostan cepat sekali di metabolisme
dan kemudian diekskresi melalui urin sehingga dapat memberikan hasil yang
berbeda bila dilakukan pada waktu yang berbeda, walaupun sampelnya sama.
2.4.3 Interleukin (IL)-6
IL-6 adalah sitokin 26 kDa yang diproduksi oleh berbagai macam sel
seperti limposit, monosit, fibroblast dan endotel termasuk juga makrofag. Melihat
spektrum fungsinya dalam proses inflamasi sistemik, IL-6 terlibat pada fase akut
dari penyakit kardiovaskular. Diproduksi bersama-sama dengan IL-1 dan TNF-
sebagai respon inflamasi di endothelium dan mungkin bekerja bersama-sama
dalam menghambat proses vasodilatasi di pembuluh darah (Woods et.al., 2000;
Wikipaedia, 2009; Davidson, 2009). Haddy etal, 2003 yang menganalisa
hubungan IL-6 dengan faktor risiko aeterosklerosis pada studi kohort
menyimpulkan bahwa dalam populasi keluarga yang sehat termasuk anak-anak,
tingkatan IL-6 berkaitan erat dengan faktor risiko aterosklerosis baik itu
43
tradisional maupun non tradisional. Sehingga data ini sangat berguna untuk
mendifinsikan peran sitokin pada mekanisme aterosklerosis pada kondisi
fisiologis. Tuomisto etal., 2006 yang diperkuat oleh Saremi etal., yang
menyatakan bahwa IL-6 adalah salah satu inflamatory marker yang berkaitan erat
dengan kejadian penyakit kardiovaskuler (Saremi A etal., 2009). Hal ini
disebabkan karena tingginya kadar IL-6 dalam serum orang sehat berkaitan erat
dengan disfungsi endotel pembuluh darah (Esteve E. etal., 2007).
Kaitannya dengan metabolisme lemak, IL-6 memiliki hubungan dengan
metabolisme lemak dimana IL-6 ini dapat menekan aktivitas dari Lipoprotein
lipase di jaringan adiposa yang mengerah kepada penekanan terhadap penyerapan
trigliserida (Greenberg etal., 1990). Hal ini akan berakibat dengan meningkatnya
kadar lemak yang kronis dalam darah yang dapat berkontribusi terhadap kejadian
aterosklerosis (Woods etal., 2000). Omoigui (2007) dalam hypotesisnya
menyebutkan bahwa IL-6 memediasi teroksidasinya LDL menjadi OxLDL dan
isoflavon ternyata dapat menekan terbentuknya IL-6. Disamping itu cholesterol
LDL yang terjebak di intima akan mengalami oksidasi menjadi oxLDL dan hal ini
akan meningkatkan ekspresi IL-6 (William and Tabbas, 1998).
Seperti disebutkan oleh Berliner et al., 1995 dan diperkuat oleh Omoigui,
2007, munculnya ROS dan jejas pada endotel ini akan merangsang terjadinya
proses inflamasi, begitu juga sebaliknya. Hal ini dapat dilihat dari banyaknya
molekul inflamasi seperti monosit adan neutrofil yang akan melekat di endotel
dan sekresi sitokin pro inflamasi. Respon yang akut diawali dengan keluarnya
sitokin proimunologik melalui aktivasi faktor transkripsi NFB (Nuclear Faktor
44
Kappa-B). Aktivasi NFB dipicu oleh berbagai stimulus, seperti sitokin inflamasi,
ROS, lemak dan kekuatan-kekuatan mekanis yang mengenai sel endotel dinding
pembuluh darah yang kemudian dapat mengaktivasi receptor trans membran
(Donato etal. 2009). Faktor ini kemudian bergerak ke dalam inti sel untuk
berikatan dengan DNA untuk mengekspresi beberapa gen target yang mengkode
sitokin-sitokin, protein fase akut dan sebagainya. Beberapa sitokin yang
diekspresi antara lain TNF-, IL1-17, IFN-. (Baraas F., 2006, ), dan terutama
yang terekpresi adalah yang dapat menyebabkan disfungsi endotil atau bersifat
proaterogenik yaitu Interleukin-6 (IL-6), Tumor Neckrosis Factor- (TNF-),
Monocyte Chemoattractant Protein 1 (MCP-1), receptor untuk Advance Glycation
End Products (RAGE) dan pro-oxidant enzyme NADPH oxidase (Donato etal.
2009). Akan tetapi ekspresi dari sitokin ini yaitu IL-6, TNF- dan CRP di dinding
pembuluh darah tidak berkaitan dengan konsentrasinya, sehingga konsentrasi
sitokin-sitokin ini saja di dalam darah tidak dapat dipakai untuk mengukur status
inflamasi di pembuluh darah (de Winther etal., 2005).
IL-6 yang terekspresi dalam sel berinti akan mudah terelminasi bersamaan
dengan lepas atau hilangnya sel tersebut di dalam darah (Melani etal., 1993).
Disamping itu produksi IL-6 menurun oleh adanya genestein yang merupakan
bagian dari flavonoid dan juga dari kalangan terpenoid (Omoigui,2007).
2.5. Antioksidan sebagai antiaterogenik
Anti oksidan didefinisikan sebagai semua substrat yang dalam jumlah yang lebih
sedikit dibanding dengan zat yang teroksidasi tapi dapat mencegah proses oksidasi
45
dari zat tersebut (Bachem et al., 1999). Sistim kerja antioksidan sangat kompleks,
tetapi secara garis besar antioksidan dapat dikelompokkan menjadi dua jenis
antioksidan yaitu: pertama, yang bersifat enzymatis, biasanya berasal dari dalam
tubuh (endogen) dan bergerak sebagai lini pertahanan pertama, antara lain:
Superoksid dismutase (SOD), Catalase (CAT), glutathione peroksidase (GPx) dan
lain-lain, dan kedua, yang bersifat radical scavenging antioxdants yang umumnya
berasal dari luar seperti: Vitamin C, B carotene, Vitamin E, flavonoid, polyphenol
dan terpenoid (Bahorun et al., 2006) (gambar 2.6). Kedua jenis antioksidan ini
bekerja bersama di dalam melawan efek dari radikal bebas (What are antioxidant,
2011).
Gambar 2.7 Diagram keterkaitan antara ROS dan antioksidan
Bahorun et al., IJMU, 1 (2), 2006
46
Walaupun dikatakan bahwa antioksidan endogen ini lebih poten daripada
yang eksogen, antioksidan dari luar ini dapat mencegah proses inisiasi dan
menterminasi berkembangnya proses oksidasi (Willcox et al., 2004).
Disebutkan bahwa ROS menyebabkan abnormalnya fungsi endotel dan
antioksidan dapat mencegahnya (Cameron and Cotter, 1999). Selain itu sel
endotel yang mengalami jejas dapat diregenerasi oleh adanya progenitor sel
endotel yang berasal dari sumsum tulang tetapi beredar di pembuluh darah, dan
ROS mengurangi efek keerja dari progenitor ini (Dimmeler and Zeiher, 2004).
2.5.1 Antioksidan Endogen
Tubuh sudah membangun sistem yang menghasilkan antioksidan yang bersifat
endogen untuk melawan Radikal Bebas. Antioksidan tersebut ada yang bersifat
enzymatik seperti SOD yang merubah O
2
-
menjadi H
2
O
2
dan H
2
O, Catalase yang
merubah H
2
O
2
menjadi H
2
O dan O
2
dan Glutathione Peroksidase yang merubah
H
2
O
2
menjadi H
2
O. Disamping itu antioksidan yang bersifat non enzymatic antara
lain Glutathione (GSH), Alpha Lipoic Acid (ALA) dan Coenzym Q10 (CoQ10)
(Fouad T, 2010, ).
Antioksidan ini disebut juga antioksidan primer karena sifatnya yang awal
yaitu mencegah munculnya radikal bebas baru dengan memutus rantai berantai
dengan mengubahnya menjadi bentuk yang lebih stabil seperti menjadi air (H
2
O)
atau hodrogen peroksida (H
2
O
2
). (Wikipedia, 2011)
NF-E2-related factor 2 (Nrf2) adalah senyawa yang berkaitan erat dengan
protein p45 NF-E2 yang bilamanan aktif dapat mengekspresi gen antioksidan
47
protein endogen. Aktifnya protein ini adalah suatu bentuk pertahanan sel
(cytoprotection) terhadap induser fase2 termasuk diantaranya stress oksidasi
(Kobayashi and Yamamoto, 2005). Senyawa yang diekspresi oleh Nrf2 antara lain
Glutathione S-transferases (GSTs) dan NAD(P)H quinone oxidoreductase (Itoh K.
etal., 1997). Disamping itu Nrf2 juga mengkode phase 2 detoxifying enzymes
seperti UDP-glucuronyl transferase 1A6, aflatoxin B1 aldehyde reductase, dan
microsomal epoxide hydrolase (Kwak etal., 2001) dan ekspresi gen enzyme
Glutamylcysteine Synthase (-GCS) yang mengkontrol sintesis GSH (Wild A.C.
etal., 1999).
2.5.2 Glutathione (GSH)
Glutathione (-glutamylcysteinylglycine, GSH) adalah antioksidan sulfhydryl (-
SH), antotoksin dan kofaktor enzym. GSH ada dimana-mana termasuk hewan,
tumbuhan, tanaman dan mikroorganisme, larut dalam air dan berada di dalam
cytosol dari sel atau substrat larut dalam air lainnya. Dan karena jumlahnya yang
cukup besar maka disebutkan sebagai antioksidan dalam sel yang mayor (Kidd P,
1997).
Glutathione eksis di dalam sel dalam bentuk antioksidan tereduksi yang
dikenal dengan istilah GSH, dan dalam bentuk teroksidasi yang dikenal dengan
istilah Glutathione Disulfida (GSSG). Rasio antara GSH/GSSG merupakan
indikator sensitif untuk stress oksidasi. GSH dengan enzym glutathione
peroksidase (GPx) dapat mengkatalisis proses reduksi Hidroperokside lemak
menjadi alkohol dan hidrogen peroksida menjadi air. Pada saat mengkatalisis tadi
ikatan disulfida dari 2 GSH akan berikatan membentuk Glutathione teroksidasi
48
(GSSG) , dan enzym glutathione reduktase dapat mendaur ulang GSSG menjadi
GSH kembali dengan cara mengoksidasi NADPH. Ketika sel terekspos dengan
stress oksidasi maka akan terjadi penumpukan GSSG dan rasio GSH/GSSG akan
menurun (Oxford Biomedical Research, 2008).
Mekanisme kerja dari GSH didalam proses peredaman radikal bebas yaitu
dalam segi kemampuananya mereduksi hidroksil radikal (
.
OH) yang berasal dari
reaksi Fenton (Best B, 2003).
Fe
++
+ H
2
O
2
> Fe
+++
+
.
OH + :OH
-
GSH +
.
OH >
.
GS + H
2
O
dan glutathione yang teroksidasi bersifat radikal akan saling menetralisir
.
GS +
.
GS > GSSG
atau dapat juga dikatakan dengan rumus yang berbeda, yaitu
2 GSH + ROOH => GSSG + ROH + H
2
O
Disamping itu enzym Glutathione peroxidase menetralisir Hidrogen
Peroksida (H
2
O
2
) dengan cara mengambil hydrogen untuk membentuk 2 H
2
O dan
satu GSSG, sedangkan enzyme glutathione reduktase akan menjadikan GSSG,
dengan menggunakan enzyme NADPH sebagai sumber hydrogen, menjadi GSH
kembali
2 GSH + H
2
O
2
=> GSSG + 2 H
2
O
Dengan kata lain glutathione di sini mencegah hidroksil radikal yang dapat
merubah molekul lemak menjadi lemak radikal (
.
L) atau peroksida lemak (LOO
.
)
melalui dua sisi yaitu mencegah terbentuknya hydroksil radikal (
.
OH) bereaksi
49
dengan molekul lemak atau mencegah terbentuknya hidroksil radikal dengan
merubah Hidrogen Peroksida (H
2
O
2
) menjadi molekul air.
Meningkatnya peroksidasi lemak dalam dinding pembuluh darah yang
mengalami aterosklerosis akan menurunkan kadar GSH peroksidase dan kadar
protector eicosanoid prostacyclin (PGI-2) mengakibatkan balans prostaglandin
menjadi lebih bersifat proinflamasi. Untuk itu diperlukan enzym GSH-S
transferase yang bekerja di sel endotel untuk meningkatkan produksi dari
protektor eicosanoid (Kidd P, 1997).
Flavonoid khususnya jenis quercetin dari ekstrak bawang bombay (onion),
kaemferol dan apigenin meningkatkan konsentrasi glutathione (GSH) melalui
aktivasi ekspresi dari -glutamylcysteine synthetase (GCS) heavy subunit (GCS
h
)
promoter (Myhrstad M.C.W. etal., 2002).
2.6 Makanan sebagai Faktor Risiko mempengaruhi Proses Penuaan
Pembuluh Darah
Gaya hidup yang meliputi makanan, alkohol, merokok dan aktivitas fisik
merupakan faktor risiko klasik terhadap penyakit pembuluh darah termasuk di
dalamnya penuaan pembuluh darah tersebut. Gaya hidup seperti diatas dianggap
dapat mempercepat ataupun mengurangi percepatan proses penuaan tersebut.
2.6.1 Makanan yang meningkatkan Risiko Menderita Penyakit Pembuluh
Darah
Li H. dan kawan-kawan (1993) menunjukkan adanya aktivasi fokal di
endotelium aorta asenden kelinci, setidaknya 1 minggu setelah pemberian diet
aterogenik yang mengandung 0,3% cholesterol, 9 % minyak kelapa yang
50
dihidrogenasi dan 1% minyak jagung dibandingkan dengan kelinci yang
mendapatkan diet 10 % minyak jagung. Bentuk fokal aktivasi yang ditunjukkan
adalah terekspresinya VCAM-1 (Vascular Cell adhesion Molecule-1) dan setelah
3 minggu di daerah fokal muncul lesi yang mengandung macrophages bearing
class II major histocomtability antigen (MHC-II) (Li H. et al., 1993). Temuan ini
ditunjang oleh Henderson dan kawan-kawan (2004) yang melakukan percobaan
pada babi, menyatakan bahwa di fase awal dari penyakit kardiovaskuler,
pemberian diet tinggi lemak dan tinggi kolesterol memberikan efek sedang
terhadap dilatasi yang dependen terhadap endotel di arteriol koroner dan efek ini
dapat dicegah bahkan dipulihkan dengan olah raga (Henderson et al., 2004). Hal
senada juga diungkapkan oleh Soinio dan kawan-kawan (2003) yang melakukan
pengamatan pada kelompok penderita Diabetes tipe 2 laki-laki dan perempuan
menyatakan bahwa pada kelompok laki-laki yang memperoleh diet lemak dengan
rasio polyunsaturated/saturated fat (P/S) tertinggi (>0,28 atau >28%) memiliki
risiko penyakit koroner terendah dibandingkan dengan yang rasio P/Snya rendah
(rasio 5 % dan 14.2 %). Dari sini Soinio menyimpulkan bahwa rendahnya rasio
P/S dapat memprediksi kemungkinan menderita penyakit jantung koroner pada
penderita Diabetes tipe 2 independen dari faktor risiko penyakit koroner
konvensional lainnya (Soinio et al., 2003). Selanjutnya Lopez-Garcia dan kawan-
kawan menyebutkan bahwa tingginya asupan trans-fatty acid secara positif
berkorelasi dengan C-reactive protein (CRP, P<0.009), soluble Tumor Necrosis
Factor Receptor-2 (sTNFR-2, P,0.002), E-selectin (P<0.003), soluble Cells
Adhesion Molecules sICAM (P,0.007), dan sVCAM (P,0.001) yang memberikan
51
efek merusak endotel dan merupakan faktor risiko dari penyakit kardiovaskuler
(Lopez-Garcia et al., 2005)
Katan dan kawan-kawan (1994) pada laporannya menyatakan bahwa
mengkonsumsi lemak jenuh akan meningkatkan kolesterol total dan LDL,
walaupun secara individu asam lemak memberikan efek yang berbeda-beda. Dari
semua jenis asam lemak jenuh, asam miristat, palmitat yang ada pada lemak
hewan dan asam laurat yang ada kelapa memberikan efek terbesar, karena asam
lemak ini yang menyebabkan LDL susah untuk dihilangkan dari sirkulasi (Hu
etal., 1999; Denke, 2006). LDL yang lama beredar dalam darah akan teroksidasi
yang nantinya akan dimakan oleh macrophage untuk menimbulkan foam cell di
tunika intima (Bahorun et al., 2006)
2.6.2 Diet yang mengandung Antioksidan dapat Mencegah Aterosklerosis
Seperti disebutkan di atas bahwa antioksidan dibedakan atas dua yaitu antioksidan
endogen yang bersifat primer dan antioksidan eksogen atau eksternal yang
umumnya berasal dari luar terutama dari makanan. Yang termasuk dalam
antitoksidan eksogen ini antara lain vitamin A, E dan C, -carotene, flavonoid dan
polyphenol dan terpenoid. Antioksidan ini biasanya berfungsi sebagai radical
scavenging antioxdants, oleh karena umumnya berasal dari luar seperti: Vitamin
C, B carotene, Vitamin E, flavonoid, polyphenol dan terpenoid (Bahorun et al.,
2006).
Disebutkan bahwa antioksidan dari luar ini dapat mencegah proses inisiasi
dan menterminasi berkembangnya proses oksidasi (Willcox et al., 2004).
52
Disamping bersifat sebagai scavenging antioxdants, flavonoid dan polypenol juga
dapat meningkatkan produksi Glutathion melalui pengaruhnya terhadap ekspresi
substrat yang diperlukan untuk sintesa glutation seperti CT cystine antiporter,
gamma-glutamylcysteine synthetase and glutathione synthase (Bahia et al., 2008),
bahkan Myhrstad et al., menyebutkan bahwa ekstrak bawang onion dan quercetin,
dapat meningkatkan konsentrasi intraseluler glutathione sampai kira-kira 50 %nya
lagi (Myhrstad et al. 2002). Vitamin E dan C terbukti telah dapat mengurangi
berkembangnya aterosklerosis. Kedua vitamin ini diketahui dapat menggagalkan
efek stimulasi Angiotensin II pada aktivitas JNK dan p38 dari Vascular Smooth
Muscle Cell (VSMC) (Kyaw et al., 2001), dan hal ini mendukung temuan dalam
penelitian yang dilakukan oleh Cambridge Heart Antioxidant Study (CHAOS)
yang dilaporkan oleh Stephens dan kawan-kawan mengatakan bahwa vitamin E
dapat menurunkan insiden Penyakit Kardiovasculer (Stephens et al., 1996).
Flavonoid oleh karena sifatnya sebagai antioksidan disebutkan dapat
mengurangi risiko menderita penyakit pembuluh darah. Antioksidan ini semakin
diperlukan dengan meningkatnya stress oksidasi oleh karena penuaan, karena
stress oksidasi bilamana dibiarkan akan merusak lapisan endotel yang berakibat
pada penyakit pembuluh darah (Achmad, 1990). Youdim dan kawan-kawan
membuat penelitian dengan komposisi polyphenol yang ada pada tanaman buah
beri dan melihat efek putatif antioksidan dan efek antiinflamasinya yang dapat
mencegah rusaknya lapisan endotel. Dari sini kelompok ini kemudian
menyimpulkan bahwa senyawa polyphenol yang ada pada buah cranberry dan
blueberry yaitu senyawa anthocyanins dan hydroxycinnamic acid dapat dideteksi
53
sampai di lapisan endotel dan dapat menurunkan kerentanan sel endotel baik di
membran maupun sampai ke sitosol. Selanjutnya senyawa polypehnol ini juga
dapat meregulasi rangsangan TNF-alpha yang diinduksi oleh mediator inflamasi
seperti IL-8, MCP-1 dan ICAM-1, sehingga senyawa ini dapat dikatakan
memberikan efek proteksi terhadap kerusakan sel endotel (Youdim et al., 2001).
Quercetin dan senyawa phenol yang ada pada jahe seperti gingerol dan shogaol
dapat mencegah penumpukan dan oksidasi LDL (Fuhrman et al., 2000, Narayana
et al., 2001). Disamping itu, zat yang ada dalam bawang putih seperti kaemferol
glicosida, merupakan scavenger yang kuat terhadap terbentuknya hydroxyl
radical dan dapat mencegah oksidasi dari LDL (Fuhrman dan Aviram, 2002).
Pendapat di atas sedikit ditentang oleh Sesso dan kawan-kawan (2003)
yang menyebutkan bahwa tidak ada hubungan linier antara asupan flavonoid
terhadap kejadian penyakit jantung koroner (Sesso et al., 2003), tetapi Xu dan
kawan-kawan (2007) menegaskan bahwa flavonoid, walaupun kemampuannnya
berbeda-beda sesuai dengan jenisnya, mempunyai efek relaksasi pembuluh darah
yang tentu sangat berguna untuk mencegah proses aterosklerosis (Xu et al. 2007)
Nevin dan Rajamohan (2003) yang melakukan penelitian tentang Virgin
Coconut Oil (VCO) menyatakan bahwa VCO menunjukkan sifat antitrombotiknya
secara signifikan di mana antioksidannya lebih tinggi, sehingga LDL lebih
resisten terhadap oksidasi dibandingkan dengan minyak bunga matahari, sehingga
disebutkan bahwa VCO dapat mencegah oksidasi dari LDL oleh oksidan, dan hal
ini oleh karena sifat VCO yang mengandung unsaponifiable component seperti
Vitamin E., provitamin A., polyphenols dan phytosterol (Nevin and Rajamohan,
54
2004). Selain itu Seneviratne et al. (2008) menyebutkan bahwa minyak kelapa
yang diolah dengan pemanasan memiliki sifat lebih stabil dibandingkan dengan
minyak kelapa yang diolah secara fermentasi
2.7. Babi Guling
Babi guling adalah salah satu makanan yang biasanya dipergunakan untuk
pelengkap kebutuhan upacara. Pada masa lalu, makanan ini biasanya dikonsumsi
oleh masyarakat Bali hanya pada waktu upacara. Tetapi makanan ini sekarang
sudah banyak diperjualbelikan di warung-warung makan, sehingga mudah untuk
dikonsumsi. Makanan ini unik karena kandungan bahan yang digunakan, disatu
sisi kandungan makanan ini, daging dan lemak, kaya akan lemak jenuh yang
bersifat aterogenik tapi disisi lain makanan ini kaya juga akan serat dan flavonoid
yang berasal dari bumbu dan tambahan lagi makanan ini juga banyak
menggunakan minyak kelapa yang belakangan ini menjadi ajang kontroversi
(Indraguna, 2009).
2.7.1 Babi Guling dalam Perpesktif Budaya
Babi guling di Bali, pada awalnya dan sampai sekarang, dikenal sebagai makanan
yang dipersembahkan pada saat upacara. Di desa adat Timbrah, bagian dari desa
Tenganan Pegringsingan dan merupakan salah satu desa adat tradisional di Bali
timur, menggunakannya sebagai salah satu kelengkapan upacara yang sampai saat
ini masih dipertahankan (Bajra, 2009). Cole (1983) dalam disertasinya tentang
prilaku makan orang bali menyebutkan bahwa babi guling merupakan salah satu
makanan upacara disamping sate, urutan, oretan, lawar, ares, tum dan sebagainya.
55
Saat ini babi guling dapat diperoleh di mana-mana. Dari restoran sampai dengan
warung lesehan kecil di seluruh kabupaten di Bali, dapat diperoleh babi guling. Di
desa Sukawati Gianyar, Babi Guling diperjual belikan dalam warung-warung
makan lesehan. Babi guling dapat dikatakan sebagai salah satu menu favorit tidak
saja di kalangan orang Bali tetapi juga mancanegara.
Bahan utama dari makanan ini adalah anak babi yang memang dipelihara
untuk itu. Untuk kebutuhan upacara, babi yang dipakai biasanya anak babi lokal
yang masih berumur muda dengan berat sekitar 8-15 kg.. Untuk kebutuhan
komersial, babi yang digunakan lebih besar yaitu beratnya mencapai 60 kg.,
bahkan dapat mencapai 80 kg. dan jenis babinyapun bukan lagi babi lokal tetapi
babi jenis Landrace atau kawin silang antara babi lokal dengan Landrace. Babi
Landrace adalah jenis babi yang awalnya dikembangkan di Denmark kemudian
diimport dan dikembangkan di Australia (Taylor et al., 2005). Babi jenis ini
sekarang sudah mulai diternakkan secara luas di Bali (Saka, 2003). Peternak babi
di Bali lebih memilih jenis babi Landrace ini karena, babi ini lebih mudah
dipelihara dibandingkan dengan babi lokal, babinya cepat besar dan dagingnya
lebih banyak sehingga lebih cepat dapat dijual dibandingkan dengan babi lokal.
Saka (1996 dan 2003) dalam laporannya menyebutkan bahwa babi Landrace ini
lebih unggul dalam memproduksikan daging karkas dibandingkan dengan babi
Saddleback maupun kawin silang antara babi bali dan babi Saddleback. Dipihak
lain pedagang babi guling khususnya yang membuat untuk tujuan komersial, juga
lebih memilih babi jenis ini karena dagingnya lebih banyak dibandingkan yang
lokal (Gung De Agung, 2009).
56
2.7.2 Pembuatan dan Penyajian Babi Guling
Babi yang akan diolah, dibunuh dengan memotong nadi di daerah leher
supaya keluar darahnya, kemudian bulu dan kulit dibersihkan dengan cara dikuliti,
dan kemudian bagian dalamnya: isi perut dan semua organ viscera dikeluarkan.
Setelah semua bersih, anus babi ditusuk dengan kayu bulat panjang melewati
perut sampai keluar dari mulut, baru kemudian rongga perut dan dada diisi dengan
bumbu. Setelah itu, perut babi dijahit kemudian kulit babi kemudian diolesi
dengan larutan dari remasan kunyit yang dicampur dengan gula merah untuk
kemudian dipanggang di atas bara api sampai dianggap matang dengan warna
kulit yang kecoklatan (gambar 2.8).
Bumbu babi guling tidak jauh berbeda dengan bumbu yang dipakai dalam
masakan tradisional Bali lainnya. Di dalam konsep bumbu Bali, bumbu
diharuskan untuk memenuhi 5 (lima) kriteria rasa yaitu rasa pahit, rasa manis, rasa
asam, rasa pedas dan rasa asin. Untuk itu komponen dalam bumbu babi guling
dibedakan menjadi tiga bagian yaitu bumbu dasar (base genep), bumbu wangn
dan bumbu penyedap (base penyangluh) (Sudharsana, 2001). Bumbu dasar terdiri
dari Lengkuas (Langua galanga), jahe (ginger atau Zingiber officinale), kencur
(Kaempferia galanga), bawang merah (Shallot, Allium Cepa), bawang putih
(garlic atau Allium Sativum), kunyit (turmeric/Alpina galanga), lombok/cab
besar (Capsicum annum), cabe rawit (bird chilli atau Capsicum frutescens),
kemiri (candle nut atau Aleurites moluccana), merica hitam (black pepper atau
Piper nigrum), ketumbar (coriander atau Coriandrum sativum) . Bumbu wangn
yang terdiri dari pala (Myristica sp.), tabya bun (Piper retrofactum), begarum,
57
cengluh dan mesui (Cinammomum), dan bumbu penyangluh yang terdiri dari
terasi (Shrimp paste), garam, daun salah ada atau tomat, daun salam (Eugenia
Polyantha), jeruk limau dan minyak kelapa (Coconut oil). Semua bumbu
dicincang halus kecuali garam, terasi, daun salam dan minyak kelapa. Pewangn
yang digunakan biasanya sudah dalam bentuk bubuk (Eiseman Jr., 1998;
Sudharsana, 2001).
Semua bumbu yang telah dicincang dicampur dengan bahan yang lain
kemudian diaduk bersama minyak kelapa yang diolah secara tradisional. Baru
kemudian semua bumbu dimasukkan ke dalam perut-dada babi yang sudah
dibersihkan.
Sebagai sayur, di beberapa daerah digunakan daun ubi kayu atau daun
kayu-manis atau daun-daunan lainnya yang berwarna hijau yang biasa dipakai
sayur, yang dicampur dengan bumbu yang kemudian dimasukkan kedalam perut
Gambar 2.8. Babi Guling Utuh
Sumber endrone.blogspot, 2009
58
babi. Di beberapa daerah, ada yang memasukkan batu hitam ke dalam perut babi
bersama-sama dengan bumbu dengan tujuan supaya daging babi matangnya
sempurna. Setelah dijahit, sebelum dipanggang kulit babi diolesi dulu dengan
minyak dan kunyit supaya kulit berwarna kecoklatan dan renyah.
Dalam penyajiannya, daging babi guling disajikan bersama dengan lemak,
kulit, sosis babai (urutan) dan gorengan organ dalam tubuh yang kemudian
dituangi bumbu diatasnya. Kemudian babi guling dikonsumsi bersama-sama
dengan nasi, lawar babi dan atau sayur nangka muda (gambar 2.9).
2.7.3 Kandungan Nutrient dan antioksidan dalam Babi Guling
Secara teoritis, daging babi mentah yang berlemak, per 100 gramnya,
mengandung 11,9 gram protein, 45 gram lemak dan 0 gram karbohidrat dan
jumlah energinya adalah 453 Kcal. Daging babi yang kurus, per 100 gramnya
Sumber endrone.blogspot, 2009
Gambar 2.9: Penyajian Babi Guling
59
mengandung 14,1 gram protein dan 35,0 gram lemak (Oey Kam Nio, 1992).
Dalam penelitian tentang inventarisasi makanan tradisional yang dilakukan oleh
Universitas Udayana Bali, disebutkan bahwa, per 100 gram daging babi guling
mengandung energi 375,76 Kkal, karbohidrat 5,96 gram, protein 8,15 gram dan
lemak 35,48 gram (Suter et al., 1999).
Tabel 2.3.
Kandungan dan Aktivitas Antioksidan dalam Bumbu Bali Guling (Secara teoritis)
FLAVONOID PHENOL TERPEN ANTIOKSIDAN
mg GAE/100 gr mVCE/100gr
Lengkuas
v 216,3* 98,61 2,13*
Kunyit
v 1340,7* v 1126,12 94,26*
Jahe
v 99,7* 62.24 0.19*
Kencur
Bawang merah
51,9* 88,9* v 11.93 2.36*
Bawang putih
63,51* 8.77 1.93*
Cabe rawit
21,91* 129,88* 130,36 2,76*
Kemiri
Ketumbar
Merica hitam
447,23*
Pala
Tabia Bun
Begarum
Cengluh
Mesui (Cinamon)
Terasi
Garam
Daun Salah Ada
Daun Salam
Jeruk limau
3,8*
Monosodium
Minyak kelapa
* Sumber: Tangkanakul etal., 2009
60
Lemak jenuh yang berasal dari babi umumnya bersifat rantai panjang
seperti palmitat (C16:0) dan stearat (C18:0) (Anne, 2008, Wikipedia, 2008, Ard
Jamy D., 2006). Seperti diketahui sebelumnya bahwa lemak jenuh rantai panjang
bersifat aterogenik, maka lemak ini dapat dikatakan sebagai makanan yang
bersifat proaging (Kromhout et al. 1995).
Beberapa bagian dari bumbu babi guling, secara individual sudah
diketahui kandungan antioksidannya, baik itu flavonoid, terpene, phenol, vitamin
A, C maupun E. Tetapi banyak juga yang belum diketahui apakah mengandung
antioksidan atau tidak. Bumbu yang sudah diketahui kandungan antioksidannya
antara lain bawang merah, lengkuas, jahe, kunyit, merica hitam, kemiri, dan jeruk
limau. Tabel 2.3 menunjukkan komposisi dari bumbu yang diambil dari
pengamatan ke beberapa produsen babi guling di Denpasar dan kandungan secara
teoritis dari masing-masing bahan yang diambil dari Tangkanakul et al., 2009.
Berdasarkan tabel 2.3, dapat dilihat bahwa komponen bumbu dalam
bumbu babi guling sangat banyak dan hampir semua bumbu seperti bawang
merah, lengkuas, bawang putih, cabe rawit dan lainnya, yang bila dilihat
kandungan antioksidan yang disumbangkan oleh masing-masing bumbu tersebut
cukup tinggi (Tzung-Hsun Tsai etal., 2005).
Berdasarkan data dari USDA Database for the Flavonoid Content of
selected Food (2003) maka disebutkan bahwa bawang merah (Allium Cepa)
mengandung quercetin, kaemferol, isorhamnetin dan cyanidin); lombok rawit
kecil mengandung quercetin, luteolin dan vitamin C; air jeruk limau selain
61
mengandung asam juga mengandung eriodyctol, naringerin dan hesperitin,
flavonols: quercetin, dan flavones: luteolin. Sedangkan kunyit mengandung
hesperitin, naringerin, eriodyctole dan quercetin. (Nutrient Data Laboratory et al.,
2003). Murtihapsari dalam percobaannya menyebutkan bahwa senyawa aktif
flavonoid tertinggi pada bawang merah adalah senyawa quercetin (Murtihapsari,
2010). Quercetin merupakan antioksidan yang kuat karena kemampuannya
bertindak sebagai scavenger terhadap radikal bebas dan ion metal. Sehingga ia
dapat menghambat peroksidasi lemak (Sakanashi etal., 2008).
Fuhrman (2002) dan Linda Lazarides (2008) dalam tulisannya menyatakan
bahwa jahe mengandung monoterpenes, sesquiterpenes, gingerol, shogaol. Verma
etal., 2004, menyebutkan bahwa jahe dapat juga mencegah peroksida lemak
disamping kemampuannya sebagai antiinflamasi (Verma etal., 2004). Benkeblia
(2005) dan Bozin et al. (2008) mengatakan kalau bawang putih dan kemiri
mengandung kaemferol, glycoside dan alysin, sedangkan Prasad etal.,
menyatakan bahwa senyawa alysin adalah suatu antioksidan yang mengikat
hydrogen peroksida (
.
OH) menjadi tidak aktif (Prasad etal., 1995). Gorenstein dan
kawan-kawan dalam penelitiannya tentang bawang putih bahkan menyebutkan
bahwa pemberian tambahan bawang putih pada tikus yang mendapatkan makanan
kaya lemak dan kolesterol, dapat menekan pembentukan plasma lipid dan
meningkatkan konsentrasi anti oksidan dalam darahnya (Gorinstein et al., 2006).
Dalam Herb and Suplement Enzyclopedia (2008) dan juga Tangkanakul
(2009) menyebutkan bahwa lengkuas mengandung galangin, gingerol,
diarylheptanoid., sedangkan Mahae N dan Chaiseri yang meneliti aktivitas
62
antioksidan secara invitro membuktikan bahwa lengkuas (Alpina Galanga) selain
mengandung ketiga unsur di atas juga mengandung methyl eugenol, chavicol,
eugenol, myricetin. Dan yang merupakan mayor antioksidan disebutkan sebagai
ekstrak 1-acetoxycavichol acetate dan catechin (Mahae and Chaiseri, 2009).
Disamping itu Aisyah Tri Septiana dan kawan-kawan (2006) menyebutkan
bahwa kandungan poli fenol pada ekstrak kunyit adalah 216,57 ppm, dan jahe
adalah 127,97 ppm. Kandungan curcuminnya untuk kunyit adalah 55,93 ppm dan
jahe adalah 4,6 ppm. Aktivitas antioksidan pada kunyit dan jahe ternyata lebih
tinggi dibandingkan dengan tocopherol. Hanumanthappa Manjunatha dan
Krishnapura Srinivasan (2008) bahkan menyebutkan bahwa kunyit dan Lombok
merah tidak hanya mengandung antioksidan tetapi juga dapat menurunkan
kolesterol darah khususnya kolesterol LDL. Tangkanakul et al., 2009, dalam
laporannya telah menghitung secara kuantitatif kandungan polyphenol dan
antioksidan yang ada pada bumbu yang dipakai masakan Thailand dan ternyata
beberapa bumbu memiliki persamaan dengan bumbu yang dipakai di Bali.
Ekstrak bawang merah dan bawang putih ternyata mampu secara
siginifikan menghambat peroksidasi lemak (Leelarungrayub et al., 2006, Yin, Shu
dan Chang, 2006), bahkan Yin, Shu dan Chang secara lebih eksplisit
menyebutkan tempat penghambatan peroksidasi lemak tersebut yaitu di
multilamellar phosphatidylcholine liposomes dan human RBC membranes.
Minyak kelapa yang diolah dengan pemanasan secara tradisional besar
kemungkinan bersifat lebih stabil dibandingkan dengan minyak kelapa yang
63
diolah secara fermentasi (Virgin Coconut Oils) (Seneviratne, 2008). Beberapa
bumbu yang lain belum diketahui kandungan antioksidannya.
Dari hasil penelitian pendahuluan yang dilakukan oleh Indraguna (2009)
diperoleh kandungan zat aktif dari bumbu adalah senyawa mengandung terpenoid,
polifenol dan flavonoid. Aktivitas antioksidan secara invitro diperoleh dapat
dilihat pada tabel 2.4.
Tabel 2.4.
Aktivitas Antioksidan pada Bumbu Babi Guling Mentah dan Matang
pada Konsentrasi yang Berbeda
KONSENTRASI
BUMBU MENTAH BUMBU MATANG
5 menit (%) 60 menit (%) 5 menit (%) 60 menit (%)
8000 ppm 109,01 116,18 131,09 251,93
1000 ppm 50,50 66,34 62,04 55,63
100 ppm 27,66 34,13 32,38 43,13
Dari tabel di atas dapat disimpulkan bahwa kemampuan bumbu sebagai
anti radikal bebas pada konsentrasi 8000 ppm mencapai di atas 100%, bahkan
dalam bumbu matang kemampuan meredam radikal bebas mencapai 250% pada
menit ke enampuluh. Pada konsentrasi 1000 ppm, kemampuan meredamnya
masih mencapai 50% ke atas, dan kemampuan bumbu matang sebagai anti radikal
bebas lebih tinggi dibandingkan dengan bumbu yang mentah (Indraguna, 2009).
Você também pode gostar
- Siklus Hidup Plasmodium Ovale Dan Plasmodium FalciparumDocumento17 páginasSiklus Hidup Plasmodium Ovale Dan Plasmodium FalciparumSheilla RatnasariAinda não há avaliações
- Vaskularisasi CerebellumDocumento1 páginaVaskularisasi CerebellumNi Made M J100% (1)
- Histologi Otot Polos Dan Otot Jantung (SP)Documento16 páginasHistologi Otot Polos Dan Otot Jantung (SP)Sisca Rizkia ArifiantiAinda não há avaliações
- Laporan Kunjungan Tawangmangu SGD 10Documento16 páginasLaporan Kunjungan Tawangmangu SGD 10rizkyanapuspita100% (1)
- Sel MerokrinDocumento6 páginasSel MerokrinSahib BroAinda não há avaliações
- Anatomi Laring SehatDocumento4 páginasAnatomi Laring Sehatlivica SanAinda não há avaliações
- Anatomi Dan Fisiologi Kelenjar KulitDocumento4 páginasAnatomi Dan Fisiologi Kelenjar KulitNatasha MengAinda não há avaliações
- Individu - PEMROSESAN JARINGANDocumento11 páginasIndividu - PEMROSESAN JARINGANAgnes Felicia LubisAinda não há avaliações
- BronkiolusDocumento3 páginasBronkiolusRahma WulanAinda não há avaliações
- Sistem Pernafasan Inspirasi Dan EkspirasiDocumento2 páginasSistem Pernafasan Inspirasi Dan EkspirasiAden Erlangga100% (1)
- Anatomi Cranium Kel 1Documento68 páginasAnatomi Cranium Kel 1Jasmine R D MAinda não há avaliações
- Anatomi KukuDocumento9 páginasAnatomi KukuTri Utami HandayaniAinda não há avaliações
- Pulvis Adspersorius (Bedak Tabur) PJJDocumento13 páginasPulvis Adspersorius (Bedak Tabur) PJJArinda Nur CahyaniAinda não há avaliações
- Potensial TransmembranpptDocumento10 páginasPotensial TransmembranpptleniAinda não há avaliações
- Mekanisme Homeostasis Dan Pembekuan Darah Melibatkan Suatu Rangkaian Proses Yang Cepat8Documento15 páginasMekanisme Homeostasis Dan Pembekuan Darah Melibatkan Suatu Rangkaian Proses Yang Cepat8Ismo YuwonoAinda não há avaliações
- Contoh Laporan Praktikum DifusiDocumento3 páginasContoh Laporan Praktikum DifusiApriliani Bambang RizkiahAinda não há avaliações
- Tugas 1 (3 Soal)Documento12 páginasTugas 1 (3 Soal)MLT 1BAinda não há avaliações
- Pengelompokan AntibiotikDocumento5 páginasPengelompokan Antibiotikartha jayantiAinda não há avaliações
- Rangkuman Imunology UasDocumento14 páginasRangkuman Imunology UasDefira ElisyiaAinda não há avaliações
- Skenario 2 OMADocumento30 páginasSkenario 2 OMAanggitekawati798Ainda não há avaliações
- Fungsi Mastikasi!Documento23 páginasFungsi Mastikasi!milhatulAinda não há avaliações
- Organ Hati Adalah Organ Yang Berperan Mengatur Homeostasis Dalam TubuhDocumento10 páginasOrgan Hati Adalah Organ Yang Berperan Mengatur Homeostasis Dalam TubuhBayu WirasmantoAinda não há avaliações
- Uji Kecepatan Pertumbuhan Rambut - KelinciDocumento16 páginasUji Kecepatan Pertumbuhan Rambut - KelinciReshy Dian PratistaAinda não há avaliações
- Makalah Struktur TulangDocumento28 páginasMakalah Struktur TulangAnggitasariAinda não há avaliações
- Jaringan EpitelDocumento53 páginasJaringan EpitelTabita PriskilaAinda não há avaliações
- PR - Reflek Pada ManusiaDocumento11 páginasPR - Reflek Pada Manusiashessy-jolycia-kerrora-3047100% (1)
- RESUME ANATOMI FISIOLOGI MANUSIA II-dikonversiDocumento4 páginasRESUME ANATOMI FISIOLOGI MANUSIA II-dikonversiSelvi RahmawatiAinda não há avaliações
- Pulvis & PulveresDocumento55 páginasPulvis & Pulvereslyna lestari indrayatiAinda não há avaliações
- Pengertian LisosomDocumento2 páginasPengertian LisosomSultanms RambeAinda não há avaliações
- Mikromeritik Again Part 2Documento23 páginasMikromeritik Again Part 2Annas RezaAinda não há avaliações
- Reseptor Pada LidahDocumento12 páginasReseptor Pada Lidahdian dayana100% (2)
- Histologi Kel. 6 MuskuloskeletalDocumento60 páginasHistologi Kel. 6 MuskuloskeletalHanna Handwi SoehandyAinda não há avaliações
- Mekanisme Menelan - Nayla 062Documento3 páginasMekanisme Menelan - Nayla 062NaylaBerlianaNugrahandhiniAinda não há avaliações
- Apoptosis Dan NekrosisDocumento7 páginasApoptosis Dan NekrosisNaditya Fitriani HasanahAinda não há avaliações
- 17 - Tca Cycle and Biological Oxidation - Luhut Uli Arto Naenggolan - 1918031020Documento28 páginas17 - Tca Cycle and Biological Oxidation - Luhut Uli Arto Naenggolan - 1918031020Luhut Arto NainggolanAinda não há avaliações
- Tahapan Siklus Sel Kontrol Pemelahan Sel Dan Mekanisme SelDocumento7 páginasTahapan Siklus Sel Kontrol Pemelahan Sel Dan Mekanisme SelRomMy AkmalAinda não há avaliações
- Arteri MaxillarisDocumento10 páginasArteri Maxillarischaerunnisa3003Ainda não há avaliações
- Refleks AchillesDocumento1 páginaRefleks AchillesChimul Lavigne 'L'Ainda não há avaliações
- Histologi Ginjal 2018Documento57 páginasHistologi Ginjal 2018theresia laurenciaAinda não há avaliações
- Anatomi MataDocumento14 páginasAnatomi MataApriliasBM100% (1)
- Cs 2009Documento76 páginasCs 2009Siti Aliyah Hani SunaryaAinda não há avaliações
- Mikroskopis Pembuluh VenaDocumento3 páginasMikroskopis Pembuluh VenaGladysouu JuaneAinda não há avaliações
- Panduan Praktikum - Bentuk Sediaan Obat - Blok 6 FKGDocumento8 páginasPanduan Praktikum - Bentuk Sediaan Obat - Blok 6 FKGJean BordeauxAinda não há avaliações
- Vena Porta HepaticaDocumento1 páginaVena Porta HepaticatasyadelizaAinda não há avaliações
- INFERTILITYDocumento30 páginasINFERTILITYPranadya ChandradewiAinda não há avaliações
- AnatomiDocumento71 páginasAnatomijulioAinda não há avaliações
- Skenario 2 KardiovaskularDocumento39 páginasSkenario 2 KardiovaskularHaya FarahAinda não há avaliações
- Vaskularisasi Jantung, Tek Darah, Pembuluh DarahDocumento15 páginasVaskularisasi Jantung, Tek Darah, Pembuluh DarahNita AndriyaniAinda não há avaliações
- Ikan Kepala Timah LaporanDocumento15 páginasIkan Kepala Timah Laporanviolita yusiAinda não há avaliações
- Pembuluh DarahDocumento11 páginasPembuluh DarahnadiavidiraAinda não há avaliações
- Syok Septi1Documento20 páginasSyok Septi1Clarithq LengguAinda não há avaliações
- Point 4& 5 FIKSDocumento18 páginasPoint 4& 5 FIKSLailatul ChamidahAinda não há avaliações
- Laporan Fishew KecebongDocumento21 páginasLaporan Fishew KecebongRioPambudiAinda não há avaliações
- Laporan Fiswan 4 - Arah Aliran Darah Pada Ikan Kepala TimahDocumento21 páginasLaporan Fiswan 4 - Arah Aliran Darah Pada Ikan Kepala TimahRizka Novitasari67% (3)
- Blood VesselsDocumento4 páginasBlood VesselsNita AndriyaniAinda não há avaliações
- Lo 4. Identifikasi Struktur Anatomi Dan Histologi Pembuluh DarahDocumento10 páginasLo 4. Identifikasi Struktur Anatomi Dan Histologi Pembuluh DarahRakha YuzartamaAinda não há avaliações
- Sistem Sirkulasi 2021 FKGDocumento9 páginasSistem Sirkulasi 2021 FKG038 Reza Zulfahmi LubisAinda não há avaliações
- Pembuluh DarahDocumento7 páginasPembuluh DarahAnastasius HendrianAinda não há avaliações
- Anatomi P. Darah Dan HistologiDocumento16 páginasAnatomi P. Darah Dan HistologiAdelAinda não há avaliações
- Aliran Darah PeriferDocumento11 páginasAliran Darah PeriferRadliya SukmajatiAinda não há avaliações
- Birth ControlDocumento6 páginasBirth ControlLioraAinda não há avaliações
- GANGGUAN KESADARAN Power PointDocumento48 páginasGANGGUAN KESADARAN Power PointNurdaily MoslemAinda não há avaliações
- DPES I - CatatanDocumento18 páginasDPES I - CatatanLioraAinda não há avaliações
- Reyer YuDocumento17 páginasReyer YuLioraAinda não há avaliações
- Lo Week TerakhirDocumento13 páginasLo Week TerakhirLioraAinda não há avaliações
- Praktikum Block Neurobehav 2017Documento4 páginasPraktikum Block Neurobehav 2017LioraAinda não há avaliações
- Praktikum Block Neurobehav 2017Documento4 páginasPraktikum Block Neurobehav 2017LioraAinda não há avaliações
- Hipertiroid HelgaDocumento7 páginasHipertiroid HelgaLioraAinda não há avaliações
- Hipotiroid HelgaDocumento4 páginasHipotiroid HelgaLioraAinda não há avaliações
- Hepatitis ADocumento18 páginasHepatitis ALioraAinda não há avaliações
- Kelenjar TiroidDocumento18 páginasKelenjar Tiroidbagus aldiAinda não há avaliações
- Wo 7,8,9Documento3 páginasWo 7,8,9LioraAinda não há avaliações
- Kelenjar TiroidDocumento18 páginasKelenjar Tiroidbagus aldiAinda não há avaliações
- Lo 3Documento2 páginasLo 3bagus aldiAinda não há avaliações
- Bahasa Indonesia JadiDocumento13 páginasBahasa Indonesia JadiLioraAinda não há avaliações
- Case Report 1 GastritisDocumento9 páginasCase Report 1 GastritisLioraAinda não há avaliações
- JGGJHGJHGDocumento2 páginasJGGJHGJHGLioraAinda não há avaliações
- Menurut GuytonDocumento5 páginasMenurut GuytonLioraAinda não há avaliações
- Diminished Urine OutputDocumento1 páginaDiminished Urine OutputLioraAinda não há avaliações
- Lo MegaDocumento4 páginasLo MegaLioraAinda não há avaliações
- Anatomi Fisiologi PankreasDocumento7 páginasAnatomi Fisiologi PankreasfadhillafianAinda não há avaliações
- CepatDocumento41 páginasCepatLioraAinda não há avaliações
- Agama 130-170Documento2 páginasAgama 130-170LioraAinda não há avaliações
- Lo AsmaDocumento13 páginasLo AsmaLioraAinda não há avaliações
- Agama 130-170Documento2 páginasAgama 130-170LioraAinda não há avaliações
- Polo PoloDocumento3 páginasPolo PoloLioraAinda não há avaliações
- Lo GuaDocumento8 páginasLo GuaLioraAinda não há avaliações
- Agama YuhuyDocumento3 páginasAgama YuhuyLioraAinda não há avaliações
- Landasan Teori IskDocumento7 páginasLandasan Teori IskLioraAinda não há avaliações
- Lapkas FaringitisDocumento9 páginasLapkas FaringitisLioraAinda não há avaliações