Você também pode gostar
- Breslin 2018Documento93 páginasBreslin 2018Rizky RayyanAinda não há avaliações
- MR Lymphangiography Reveals Central Lymphatic PathwaysDocumento14 páginasMR Lymphangiography Reveals Central Lymphatic PathwaysHector Hernandez-SoriaAinda não há avaliações
- Kejang NeonatusDocumento12 páginasKejang NeonatustriAinda não há avaliações
- Lymphotropic Therapy For Diseases of The Maxillofacial Region Review of The LiteratureDocumento6 páginasLymphotropic Therapy For Diseases of The Maxillofacial Region Review of The LiteratureEditor IJTSRDAinda não há avaliações
- The lymphatic system's role in collecting and transporting tissue fluidsDocumento3 páginasThe lymphatic system's role in collecting and transporting tissue fluidsTania Louise Pioquinto AbuanAinda não há avaliações
- Fisiologia LinfedemaDocumento8 páginasFisiologia LinfedemaSAKAI69Ainda não há avaliações
- 15 Lymphatic SystemDocumento4 páginas15 Lymphatic SystemOribello, Athenna Jae W.Ainda não há avaliações
- Jennings 2008 ImagingHemodynamicsDocumento25 páginasJennings 2008 ImagingHemodynamicsAhmed El KaffasAinda não há avaliações
- Lymph SystemDocumento7 páginasLymph SystemghaiathAinda não há avaliações
- Sample Essay BiologyDocumento7 páginasSample Essay Biologyapi-336421734Ainda não há avaliações
- The Lymphatics in Kidney Health and Disease: Michael D. Donnan, Yael Kenig-Kozlovsky and Susan E. QuagginDocumento21 páginasThe Lymphatics in Kidney Health and Disease: Michael D. Donnan, Yael Kenig-Kozlovsky and Susan E. QuagginsilviaAinda não há avaliações
- Cardiovascular LymphaticDocumento17 páginasCardiovascular LymphaticFaisal Ridho SaktiAinda não há avaliações
- Department of Histology, Cytology and EmbryologyDocumento8 páginasDepartment of Histology, Cytology and Embryologysubmen81Ainda não há avaliações
- Anatomyand Physiologyofthe Thoraciclymphatic SystemDocumento15 páginasAnatomyand Physiologyofthe Thoraciclymphatic SystemLis Borda MuñozAinda não há avaliações
- Lec. 1 Selected TopicsDocumento30 páginasLec. 1 Selected TopicsMustafa KamelAinda não há avaliações
- Modeling and Analysis of Blood Flow in Human Circulatory SystemDocumento3 páginasModeling and Analysis of Blood Flow in Human Circulatory SystemPradeep K SinghAinda não há avaliações
- Anatomy of Lymphatics SystemDocumento9 páginasAnatomy of Lymphatics Systemtg856qr4mwAinda não há avaliações
- 24circulation Part 2Documento11 páginas24circulation Part 2Jaydave PatelAinda não há avaliações
- Chapter 16Documento9 páginasChapter 16g_komolafeAinda não há avaliações
- The Lymphatic SystemDocumento7 páginasThe Lymphatic SystemPreeti ChouhanAinda não há avaliações
- Microcirculation & Lymphatics - New-1Documento23 páginasMicrocirculation & Lymphatics - New-1Danial HaiderAinda não há avaliações
- TanisDocumento11 páginasTanisZULMA MADELINE LIBNY GUTARRA TICAAinda não há avaliações
- Anatomy of The Lymphatic SystemDocumento27 páginasAnatomy of The Lymphatic SystemAntonio Lima Filho100% (2)
- Tumor Cell Interaction with Lymphatic Vessels in Cancer Progression and MetastasisDocumento10 páginasTumor Cell Interaction with Lymphatic Vessels in Cancer Progression and MetastasisChetan PatilAinda não há avaliações
- Anatomy of The Lymphatic and Immune Systems - NoteDocumento18 páginasAnatomy of The Lymphatic and Immune Systems - NoteRashini FernandoAinda não há avaliações
- Anatomy and Physiology MidtermsDocumento39 páginasAnatomy and Physiology MidtermsJingky AntonioAinda não há avaliações
- Physical Review Letters Volume 117 issue 13 2016 [doi 10.1103%2FPhysRevLett.117.138301] Ronellenfitsch, Henrik_ Katifori, Eleni -- Global Optimization, Local Adaptation, and the Role of Growth in Dist (1).pdfDocumento5 páginasPhysical Review Letters Volume 117 issue 13 2016 [doi 10.1103%2FPhysRevLett.117.138301] Ronellenfitsch, Henrik_ Katifori, Eleni -- Global Optimization, Local Adaptation, and the Role of Growth in Dist (1).pdfMichele HosackAinda não há avaliações
- Lymphoid Organs ReviewDocumento9 páginasLymphoid Organs Reviewshko noshaAinda não há avaliações
- Chapter 8 - Circulation and ImmunityDocumento94 páginasChapter 8 - Circulation and Immunitychloe06muller.mAinda não há avaliações
- Diuretics: Dr. Ashley M. StokesDocumento26 páginasDiuretics: Dr. Ashley M. StokesDhaif dhaifAinda não há avaliações
- Gashev 2006Documento15 páginasGashev 2006yalocim666Ainda não há avaliações
- tjp0540 1023Documento15 páginastjp0540 1023hjakamjAinda não há avaliações
- Yearly Plan Bio Form5Documento30 páginasYearly Plan Bio Form5maieyAinda não há avaliações
- Comparing Transport in Plants & AnimalsDocumento2 páginasComparing Transport in Plants & AnimalsBio SciencesAinda não há avaliações
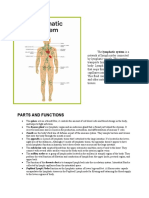
- Parts and FunctionsDocumento4 páginasParts and FunctionsGerwin SilvestreAinda não há avaliações
- Notes 20Documento3 páginasNotes 20Paris ShaShaAinda não há avaliações
- Lymphatic System Stu.Documento4 páginasLymphatic System Stu.Rinopro 555Ainda não há avaliações
- 4 Circulatory SystemDocumento23 páginas4 Circulatory SystemSundari D'yojapa NonemAinda não há avaliações
- Lymphatic System: Roles, Functions, and Differences from Blood CirculationDocumento18 páginasLymphatic System: Roles, Functions, and Differences from Blood CirculationNorJannah RAinda não há avaliações
- The Anatomy of The Immune System (Lymphatic SystemDocumento4 páginasThe Anatomy of The Immune System (Lymphatic SystemMohammed R.HusseinAinda não há avaliações
- Lymphatic System ReportDocumento6 páginasLymphatic System ReportcrtgyhujikAinda não há avaliações
- Lymphatic System and Immunity ChapterDocumento18 páginasLymphatic System and Immunity ChapterElvin MoletaAinda não há avaliações
- Nihms 1585884Documento17 páginasNihms 1585884CT DAMAinda não há avaliações
- Cardiovascular and Respiratory Lecture Handout 2023-24Documento40 páginasCardiovascular and Respiratory Lecture Handout 2023-24extreme fattypunchAinda não há avaliações
- Lymphatic SystemDocumento70 páginasLymphatic SystemNang Maizana Megat Yahya100% (1)
- Llymphatic SystemDocumento14 páginasLlymphatic SystemfauziharunAinda não há avaliações
- Anatomy and Physiology of The Circulatory and Ventilatory Systems, ThirietDocumento594 páginasAnatomy and Physiology of The Circulatory and Ventilatory Systems, ThirietSubhajit Chakraborty100% (1)
- Lymphatic SystemDocumento5 páginasLymphatic SystemEmmanuel A. GalvanAinda não há avaliações
- (Robert H Harris) Textbook of Dr. Vodder's ManualDocumento116 páginas(Robert H Harris) Textbook of Dr. Vodder's Manualmitroisergiu100% (10)
- Lymphatic SystemDocumento28 páginasLymphatic SystemRozi Reviana PratiwiAinda não há avaliações
- Anatomy of the Lymphatic and Immune Systems ExplainedDocumento8 páginasAnatomy of the Lymphatic and Immune Systems ExplainedAbdallah AlasalAinda não há avaliações
- Regulation of Tissue Oxygenation: Second EditionDocumento114 páginasRegulation of Tissue Oxygenation: Second EditionrezqAinda não há avaliações
- PLEURAL EFFUSIONS CAUSES AND DIAGNOSISDocumento5 páginasPLEURAL EFFUSIONS CAUSES AND DIAGNOSISnfacmaAinda não há avaliações
- Lymphatic SystemDocumento13 páginasLymphatic SystemCharli ParachinniAinda não há avaliações
- The Lymphatic Fluid: Formation, Circulation, and Immunological RoleDocumento23 páginasThe Lymphatic Fluid: Formation, Circulation, and Immunological RolektorooAinda não há avaliações
- Fisiopatologia Linfedema 1997Documento6 páginasFisiopatologia Linfedema 1997ktorooAinda não há avaliações
- Intrahepatic Cholangiocarcinoma Surgery The ImpactDocumento10 páginasIntrahepatic Cholangiocarcinoma Surgery The Impactraplyhollow_68780740Ainda não há avaliações
- Anatomy of The Human Lymphatic System: Department of Surgery, University of Sao Paulo, BrazilDocumento22 páginasAnatomy of The Human Lymphatic System: Department of Surgery, University of Sao Paulo, BrazilAprodita Permata YulianaAinda não há avaliações
- Importance of lymph in human balanceDocumento1 páginaImportance of lymph in human balanceZaid Bin salmanAinda não há avaliações
- Computational Models in Biomedical Engineering: Finite Element Models Based on Smeared Physical Fields: Theory, Solutions, and SoftwareNo EverandComputational Models in Biomedical Engineering: Finite Element Models Based on Smeared Physical Fields: Theory, Solutions, and SoftwareAinda não há avaliações
- Semiologia UrológicaDocumento137 páginasSemiologia UrológicaAntonio Fernandes Neto100% (3)
- HelloDocumento1 páginaHellonnimrodoAinda não há avaliações
- HelloDocumento1 páginaHellonnimrodoAinda não há avaliações
- HelloDocumento1 páginaHellonnimrodoAinda não há avaliações
- Anhidrosis in EquineDocumento3 páginasAnhidrosis in EquineZain AsifAinda não há avaliações
- Language and BrainDocumento23 páginasLanguage and Brainalbino100% (1)
- Bac ToxinDocumento9 páginasBac ToxinJawairia KhawarAinda não há avaliações
- Nursing Care Plans for Acute Pain, Hyperthermia, Activity Intolerance, and Infection RiskDocumento20 páginasNursing Care Plans for Acute Pain, Hyperthermia, Activity Intolerance, and Infection RiskNursidar Pascual Mukattil80% (5)
- Bio 320 Practical 7Documento12 páginasBio 320 Practical 7MuhammadAsyraf50% (2)
- Enteral NutritionDocumento11 páginasEnteral NutritionEso Soso0% (1)
- Motility Disorders of EsophagusDocumento44 páginasMotility Disorders of EsophagusmackieccAinda não há avaliações
- Zoology Practical Exam Tips 2024Documento62 páginasZoology Practical Exam Tips 2024sureshsooraj426Ainda não há avaliações
- Neuroparasitology PDFDocumento62 páginasNeuroparasitology PDFpsetoAinda não há avaliações
- Snakes in India - Tradition and The Truth PDFDocumento5 páginasSnakes in India - Tradition and The Truth PDFhoticeforuAinda não há avaliações
- 1 - OB 1 (Maternal Anatomy)Documento13 páginas1 - OB 1 (Maternal Anatomy)Gen XAinda não há avaliações
- A Presentation On Pantothenic Acid or b5Documento28 páginasA Presentation On Pantothenic Acid or b5jainsaketAinda não há avaliações
- Throwing RehabDocumento16 páginasThrowing RehabSean GlynnAinda não há avaliações
- Assessment of Fluid and ElectrolyteDocumento15 páginasAssessment of Fluid and ElectrolyteDip Ayan MAinda não há avaliações
- Health Benefits of Running and JoggingDocumento2 páginasHealth Benefits of Running and JoggingOsamaAinda não há avaliações
- Pregnant woman with painful swollen legDocumento12 páginasPregnant woman with painful swollen legjimmojonesAinda não há avaliações
- Hyperosmolar Hyperglycemic Nonketotic Syndrome: Presented by DR MoosallyDocumento10 páginasHyperosmolar Hyperglycemic Nonketotic Syndrome: Presented by DR MoosallyDrsarfaraz SiddiquiAinda não há avaliações
- Brewing Technology (2017)Documento190 páginasBrewing Technology (2017)Јован АдамовAinda não há avaliações
- Yellow Belt Level 01 - Solution - 0Documento2 páginasYellow Belt Level 01 - Solution - 0Jo Marie LeeAinda não há avaliações
- The Ionization of Amino Acids at Physiological pHDocumento4 páginasThe Ionization of Amino Acids at Physiological pHNatasha MahatantilaAinda não há avaliações
- Fluid Volume DeficitDocumento2 páginasFluid Volume DeficitpeternohibiAinda não há avaliações
- NCPDocumento10 páginasNCPbabycheska08Ainda não há avaliações
- Newborn Care PDFDocumento8 páginasNewborn Care PDFZedrake CaraanAinda não há avaliações
- Analgesia AvesDocumento16 páginasAnalgesia AvesJESSICA ZAPATAAinda não há avaliações
- Pediatric DehydrationDocumento27 páginasPediatric DehydrationAbraham ChiuAinda não há avaliações
- ANS & CVS Response To Exercise v2Documento18 páginasANS & CVS Response To Exercise v2chow wing yin amandaAinda não há avaliações
- Prokaryotic Cells: Eukaryotes Vs Prokaryotes: What's The Difference?Documento8 páginasProkaryotic Cells: Eukaryotes Vs Prokaryotes: What's The Difference?Anggun Teh PamegetAinda não há avaliações
- Week 006 Animal-Immune-SystemDocumento13 páginasWeek 006 Animal-Immune-SystemMark Lorens StaanaAinda não há avaliações
- Liver Cirrhosis NCP 1Documento6 páginasLiver Cirrhosis NCP 1Paolo Rafael D EsguerraAinda não há avaliações
- Change Your Life NowDocumento18 páginasChange Your Life NowAbdul Lateef Khan0% (1)


























![Physical Review Letters Volume 117 issue 13 2016 [doi 10.1103%2FPhysRevLett.117.138301] Ronellenfitsch, Henrik_ Katifori, Eleni -- Global Optimization, Local Adaptation, and the Role of Growth in Dist (1).pdf](https://imgv2-1-f.scribdassets.com/img/document/335024555/149x198/c2af002bd6/1482684538?v=1)