Escolar Documentos
Profissional Documentos
Cultura Documentos
Biologia
Enviado por
enzosp180 notas0% acharam este documento útil (0 voto)
25 visualizações12 páginashola
Título original
biologia
Direitos autorais
© © All Rights Reserved
Formatos disponíveis
DOCX, PDF, TXT ou leia online no Scribd
Compartilhar este documento
Compartilhar ou incorporar documento
Você considera este documento útil?
Este conteúdo é inapropriado?
Denunciar este documentohola
Direitos autorais:
© All Rights Reserved
Formatos disponíveis
Baixe no formato DOCX, PDF, TXT ou leia online no Scribd
0 notas0% acharam este documento útil (0 voto)
25 visualizações12 páginasBiologia
Enviado por
enzosp18hola
Direitos autorais:
© All Rights Reserved
Formatos disponíveis
Baixe no formato DOCX, PDF, TXT ou leia online no Scribd
Você está na página 1de 12
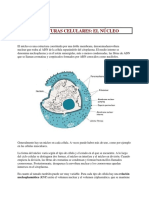
Sistema endomembranoso.
El sistema endomembranoso es el sistema de membranas internas de las
clulas eucariotas que divide la clula en compartimientos funcionales y
estructurales, denominados orgnulos. Las procariotas no tienen un
sistema endomembranoso y as carecen de la mayora de los orgnulos.
El sistema endomembranoso tambin proporciona un sistema de
transporte para las molculas mviles a travs del interior de la clula, as
como superficies interactivas para la sntesis de lpidos y de protenas. Las
membranas que componen el sistema endomembranosos se construyen a
partir de una bicapa lpida, con las protenas unidas a cada lado o
atravesndolas.-
Los orgnulos siguientes son parte del sistema endomembranoso:
_El retculo endoplasmtico es un orgnulo de sntesis y transporte
construido como una extensin de la membrana nuclear.
_El aparato de Golgi acta como el sistema de empaquetado y de entrega
de molculas.
_Los lisosomas son las unidades energeticas de la clula. Utilizan
enzimas que analizan las macromolculas y tambin actan como sistema
de recogida de residuos.
_Las vacuolas actan como unidades del almacenaje en algunas clulas.
_Las vesculas son pequeas unidades de transporte delimitadas por
membranas que pueden transferir molculas entre diversos
compartimientos.
Desarrollo y formacin del sistema de endomembranas
Este sistema comienza el retculo endoplasmtico rugoso (RER) que es un
conjunto de pliegues membranosos que son prolongacin de la
membrana celular. En l la principal funcin es la sntesis de protenas. De
ellas algunas se almacenan para luego llevarlas a otros orgnulos; y otras
se quedan adosadas al mismo para hacer crecer sus membranas. Llegado
a un punto el RER pierde los ribosomas y forma tbulos y cisternas
membranosas donde principalmente se forman lpidos: el retculo
endoplasmtico liso. En l se generan unas glndulas que secretan
vesculas de transicin que se van uniendo formando el dictiosoma con
forma de media luna. El dictiosoma es la unidad de funcionamiento del
aparato de Golgi. El conjunto de dictiosomas de una clula forma el
aparato de Golgi. Esta estructura finaliza con unas vesculas de sntesis
que son secretadas al citoplasma y luego darn lugar a los lisosomas,
peroxisomas y vacuolas.
EL RETICULO ENDOPLASMATICO
El retculo endoplasmtico es un complejo sistema de membranas
dispuestas en forma de sacos aplanados y tbulos que estn
interconectados entre s compartiendo el mismo espacio interno. Sus
membranas se continan con las de la envuelta nuclear y se pueden
extender hasta las proximidades de la membrana plasmtica, llegando a
representar ms de la mitad de las membranas de una clula. Debido a
que los cidos grasos que las componen suelen ser ms cortos son ms
delgadas que las dems. El retculo organiza sus membranas en regiones o
dominios que realizan diferentes funciones. Los dos dominios ms fciles
de distinguir son el retculo endoplasmtico rugoso, con sus membranas
formando tbulos ms o menos rectos, a veces cisternas aplanadas, y con
numerosos ribosomas asociados, y el retculo endoplasmtico liso, sin
ribosomas asociados y con membranas organizadas formando tbulos
muy curvados e irregulares. La membrana externa de la envuelta nuclear
se puede considerar como parte del retculo endoplasmtico puesto que
es una continuacin fsica de l y se pueden observar ribosomas asociados
a ella realizando la traduccin. El retculo endoplasmtico rugoso y el liso
suelen ocupar espacios celulares diferentes como ocurre en los
hepatocitos, en las neuronas y en las clulas que sintetizan esteroides. Sin
embargo, en algunas regiones del retculo no existe una segregacin clara
entre ambos dominios y se aprecian reas de membrana con ribosomas
mezcladas con otras sin ribosomas. La disposicin espacial del retculo
endoplasmtico en las clulas animales depende de sus interacciones con
los microtbulos, mientras que en las vegetales son los filamentos de
actina los responsables.
*El retculo endoplasmtico es un orgnulo cuyas membranas forman
cisternas aplanadas y tbulos conectados entre s. Posee dos dominios
morfolgicos y funcionales: rugoso, cisternas aplanadas con ribosomas
asociados a sus membranas, y liso, tbulos.
En el retculo endoplasmtico rugoso se sintetizan protenas para su
secrecin o para formar parte de otros compartimentos membranosos
que participan de la ruta vesicular.
En el retculo endoplasmtico liso se sintetizan numeroso lpidos de las
membranas, es un centro metablico de detoxificacin, almacn de
Retculo endoplasmtico rugoso
RETICULO ENDOPLASMATICO RUGOSO
El dominio rugoso del retculo endoplasmtico se caracteriza por
organizarse en una trama de tbulos alargados o sacos aplanados y
apilados, ms o menos regulares en su forma, con numerosos ribosomas
asociados a sus membranas. La cantidad de ribosomas asociados a sus
membranas condiciona la forma de este orgnulo, de tal manera que
cuando el nmero de ribosomas asociados aumenta los tbulos se
expanden adoptando la forma de cisternas aplanadas. Esta morfologa es
claramente visible en imgenes tomadas con el microscopio electrnico
de las clulas secretoras de protenas, las cuales tienen el retculo
endoplasmtico muy desarrollado.
La principal misin del retculo endoplasmtico rugoso es la sntesis de
protenas que irn destinadas a diferentes lugares: el exterior celular, el
interior de otros orgnulos que participan en la ruta vesicular, como los
lisosomas, o que formarn parte integral de las membranas, tanto
plasmtica como de otros orgnulos de la ruta vesicular. Adems, el
retculo endoplasmtico rugoso tiene que sintetizar protenas para s
mismo, denominadas protenas residentes. Las protenas integrales de la
membrana plasmtica se sintetizan en el retculo endoplasmtico. Hay
que tener en cuenta que las protenas sintetizadas en el retculo que van
destinadas a orgnulos concretos de la ruta vesicular deben tener unas
secuencias de aminoidos o modificaciones especficas (actuarn como
seales) para que cuando lleguen a la zona de reparto del aparato de
Golgi sean reconocidas y dirigidas a sus compartimentos diana
correspondientes.
Cualquier protena que se secrete o que forme parte de los orgnulos o
compartimentos de la ruta vesicular empieza su proceso de sntesis en el
citosol pero terminar en el interior de una cisterna del retculo o
formando parte de sus membranas. El proceso comienza con la unin de
los ARNm, localizados en el citosol, unindose en primer lugar a una
subunidad pequea ribosomal y posteriormente a una subunidad grande
ribosomal para comenzar la traduccin. Lo primero que se traduce de
estos ARNm es una secuencia inicial de nucletidos a partir de la cual se
sintetiza una cadena de unos 70 aminocidos denominada pptido seal.
Una molcula conocida como SRP (sequence recognizing particule), que
es una mezcla de 7 ARNs y 6 polipptidos, reconoce al pptido seal y
enlentece el proceso de traduccin. El complejo formado por ribosoma,
ARNm, pptido seal ms SRP difunde por el citosol hasta chocar con las
membranas del retculo endoplasmtico, a las cuales se une gracias a la
existencia de un receptor de membrana que reconoce al SRP. Todo el
complejo anterior interacciona con un translocador, que es una protena
integral que forma un canal por el cual penetra la cadena polipeptdica
naciente hacia el interior de la cisterna del retculo endoplasmtico. El
pptido seal queda unido al translocador mientras que el resto de la
cadena que se va traduciendo y liberando hacia el interior. Una peptidasa
presente en el retculo escinde el pptido seal del resto de la cadena de
aminocidos, quedando sta libre en el interior. Una vez completada la
sntesis, la cadena de aminocidos adopta su conformacin tridimensional
y el ribosoma se libera de la membrana.
Si la protena que se est sintetizando es una protena integral de
membrana no ser liberada al interior del retculo. En estos casos las
protenas nacientes tienen cadenas de aminocidos hidrfobos que
cuando se traducen facilitan su insercin directamente entre los cidos
grasos de la membrana gracias a la accin del translocador. El proceso es
muy complejo y diverso para los diferentes tipos de protenas integrales
puesto que en un receptor con siete cruces de membrana tienen que
alternarse secuencias que han de insertarse en la membrana con otras
que quedan bien en el lado citoslico o bien en el interior de la cisterna
del retculo endoplasmtico. Slo en raras ocasiones el retculo importa
protenas que se sintetizan completamente en el citosol gracias a ciertos
transportadores presentes en su membrana.
Las protenas que se sintetizan en los ribosomas adosados a la membrana
del retculo endoplasmtico son modificadas conforme van siendo
sintetizadas. a) Hay una glucosilacin (N-glucosilacin) de los aminocidos
asparragina. stos recibirn un complejo de 14 azcares en su radical, que
son transferidos desde un lpido embebido en la membrana denominado
dolicol fosfato, perdindose algunos de estos azcares en procesos
posteriores. b) Se da hidroxilacin slo en algunas protenas, sobre todo
en aquellas que van a formar parte de la matriz extracelular. Aqu se
hidroxilan los aminocidos prolina y lisina, dando hidroxiprolina e
hidroxilisina, que formarn parte del colgeno. c) Algunas protenas
asociadas a la membrana plasmtica estn unidas covalentemente a
lpidos de la membrana, esta unin tambin se produce en este
compartimento.
En el retculo endoplasmtico se produce un control de la calidad de las
protenas sintetizadas, de modo que aquellas que tienen defectos son
sacadas al citosol y eliminadas. Existen unas protenas denominadas
chaperonas que juegan un papel esencial en el plegamiento y maduracin
de las protenas recin sintetizadas. Son tambin ellas las encargadas de
detectar errores y marcar las protenas defectuosas para su degradacin.
Otras protenas con dominios tipo lectina, reconocen determinados
azcares y comprueban la adicin correcta de glcidos.
Vesculas
Formacin y fusin de vesculas.
Como dijimos, las protenas que se sintetizan en el retculo
endoplasmtico terminan en varios posibles destinos: en el exterior
celular mediante un proceso de secrecin, el interior o en la membrana
de alguno de los compartimentos de la ruta vesicular como el aparato de
Golgi, los endosomas o los lisosomas. Sin embargo, algunas tienen su
funcin en el propio retculo endoplasmtico, son las denominadas
protenas residentes. Hemos nombrado algunas como las chaperonas,
ciertas glusosidasas, el receptor para el SRP, el propio translocados,
etctera. Para ser retenidas en el retculo deben poseer una secuencia de
cuatro aminocidos concrfetos localizados en el extremo carboxilo (-
COOH).
Retculo endoplasmtico liso
Es un entramado de tbulos membranosos interconectados entre s y que
se continan con las cisternas del retculo endoplasmtico rugoso. No
tienen ribosomas asociados a sus membranas, de ah el nombre de liso,
por tanto la mayora de las protenas que contiene son sintetizadas en el
retculo endoplasmtico rugoso. Es abundante en aquellas clulas
implicadas en el metabolismo de grasas, detoxificacin y almacn de
calcio.El retculo endoplasmtico liso est involucrado en una serie de
importantes procesos celulares de los que se pueden destacar:
Sntesis lipdica
Las membranas del retculo endoplasmtico liso producen la mayora de
los lpidos requeridos para la elaboracin de las nuevas membranas de la
clula, incluyendo glicerofosfolpidos y colesterol. Gran parte de la sntesis
de los esfingolpidos se lleva a cabo en el aparato de Golgi, pero su
estructura bsica, la ceramida, se sintetiza tambin en el retculo. En
realidad, en las membranas del retculo no se realizan todos los pasos de
la sntesis de los lpidos de las membranas. Los cidos grasos se sintetizan
en el citosol y son insertados posteriormente en las membranas del
retculo endoplasmtico liso donde son transformados en
glicerofosfolpidos. Esta sntesis la realizan protenas de membrana que
tienen sus centros activos orientados hacia el citosol y por tanto los
lpidos estarn inicialmente en la hemicapa citoslica de la membrana.
Como el cambio pasivo de los lpidos entre hemicapas, o movimiento
"flip-flop", es difcil por el ambiente hidrfobo de las cadenas de cidos
grasos de la membrana, para que algunos de ellos lleguen a la hemicapa
interna desde la externa se requiere la existencia de transportadores de
lpidos. Hay varios tipos de transportadores de ldos entre las dos
hemicapas de la membrana que se distribuyen por los diferentes
compartiementos membranosos, includa la membrana plasmtica. Unos
estn especializados en la transportar desde la hemicapa citoslica hasta
la extracelular (el espacio interno de los orgnulos se puede considerar
como extracelular), otros lo hacen en sentido contrario. A stas se las
denomina flipasas y flopasas, respectivamente, y consumen energa. Hay
otras denominadas "mezcladoras" (scramblases en ingls) que
transportan lpidos en ambas direcciones. Por tanto, la asimetra se
establece a lo largo de la va vesicular y las caractersticas de esta
asimetra depende de las protenas transportadoras que posean. Tambin
contribuyen a la asimetr os azcares que formarn los glucolpidos se
ensamblan en el interior del retculo y del aparato de Golgi, y no sern
expuestos al citosol. Por tanto, en cuestin de lpidos, la localizacin
asimtrica de stos en las membranas no depende de la sntesis inicial en
el retculo endoplasmtico.
Transportador
Esquema de los caminos propuestos para el transporte de lpidos desde el
retculo endoplasmtico hasta otras membranas celulares: en vesculas y
mediante transportadores.
El colesterol es otro importante componente de las membranas, sobre
todo de la plasmtica, que se sintetiza mayoritariamente en el retculo
endoplasmtico liso. Desde aqu es transportado por la va vesicular o por
transportadores proteicos solubles. Por ejemplo, las levaduras, que
poseen ergosterol en sus membranas en vez de colesterol, usan vas no
vesiculares para transportar el ergosterol desde el retculo hasta la
membrana plasmtica. Estos transportadores son diversos y sus
movimientos son independientes de ATP.
Las mitocondrias y los peroxisomas no forman parte de la ruta vesicular
por lo que sus lpidos de membrana deben ser importados. Para ello
utilizan los transportadores de lpidos. Por ejemplo, para los
glicerofosfolpidos existen unas protenas solubles llamadas
intercambiadoras de glicerofosfolpidos que tienen la habilidad de
transportarlos a travs del citosol. Los toman en la membrana del retculo
endoplasmtico liso y los sueltan en las de estos orgnulos. En las clulas
de los tejidos fotosintticos son los cloroplastos los encargados de
sintetizar sus propios glicerofosfolpidos y glucolpidos.
En el retculo endoplasmtico liso tambin se sintetizan lpidos como los
triacilgliceroles que sern almacenados en el propio retculo. Este proceso
es muy activo en los adipocitos, clulas que almacenan grasa, con dos
funciones: reserva alimenticia y aislamiento trmico. Tambin es el
principal responsable de la sntesis de la parte lipdica de las lipoprotenas,
de la produccin de hormonas esteroideas y de cidos biliares.
*
Detoxificacin
Los hepatocitos, las clulas tpicas del hgado, tienen un retculo
endoplasmtico liso muy desarrollado. En l se sintetizan las lipoprotenas
que transportarn al colesterol y a otros lpidos al resto del organismo. En
sus membranas se encuentran tambin enzimas, como la familia de
protenas P450, responsables de la eliminacin de productos del
metabolito potencialmente txicos, as como algunas toxinas liposolubles
incorporadas durante la ingesta. La superficie de membrana del retculo
se adapta a la cantidad de enzimas detoxificadoras sintetizadas, la cual
depende a su vez de la cantidad de txicos presentes en el organismo.
Defosforilacin de la glucosa-6 fosfato.
La glucosa se suele almacenar en forma de glucgeno, fundamentalmente
en el hgado. Este rgano es el principal encargado de aportar glucosa a la
sangre, gracias a la regulacin llevada a cabo por las hormonas glucagn e
insulina. La degradacin del glucgeno produce glucosa-6-fosfato que no
puede atravesar las membranas y por tanto no puede abandonar las
clulas. La glucosa 6-fosfatasa se encarga de eliminar ese residuo fosfato,
permitiendo que la glucosa sea transportada al exterior celular.
Reservorio intracelular de calcio
Las cisternas del retculo endoplasmtico liso estn tambin
especializadas en el secuestro y almacenaje de calcio procedente del
citosol. Este calcio puede salir de forma masiva en respuesta a seales
extra o intracelulares gracias a cascadas de segundos mensajeros. Un
ejemplo destacable es el retculo sarcoplsmico (nombre que recibe el
retculo endoplasmtico liso en las clulas musculares) que secuestra
calcio gracias a una bomba de calcio presente en sus membranas. El
secuestro y la salida de calcio desde el retculo sarcoplsmico se produce
en cada ciclo de contraccin de la clula musculacalcio, etctera
EL APARATO DE GOLGI
El aparato de Golgi es un orgnulo presente en todas las clulas
eucariotas. Pertenece al sistema de endomembranas. Est formado por
unos 80 dictiosomas (dependiendo del tipo de clula), y estos dictiosomas
estn compuestos por 40 o 60 cisternas (sculos) aplanadas rodeados de
membrana que se encuentran apilados unos encima de otros, y cuya
funcin es completar la fabricacin de algunas protenas. Funciona como
una planta empaquetadora, modificando vesculas del retculo
endoplasmtico rugoso. El material nuevo de las membranas se forma en
varias cisternas del aparato de Golgi. Dentro de las funciones que posee el
aparato de Golgi se encuentran la glicosilacin de protenas, seleccin,
destinacin, glicosilacin de lpidos, almacenamiento y distribucin de
lisosomas, al igual que los peroxisomas, que son vesculas de secrecin de
sustancias. La sntesis de polisacridos de la matriz extracelular. Debe su
nombre a Camillo Golgi, Premio Nobel de Medicina en 1906 junto a
Estructura del aparato de Golgi
El aparato de Golgi se compone en estructuras denominadas sculos.
stas se agrupan en nmero variable, habitualmente de 4 a 8, formando
el dictiosoma en las plantas. Presentan conexiones tubulares que
permiten el paso de sustancias entre las cisternas. Los sculos son
aplanados y curvados, con su cara convexa (externa) orientada hacia el
retculo endoplasmtico. Normalmente se observan entre 4 y 8, pero se
han llegado a observar hasta 60 dictiosomas.1 Alrededor de la cisterna
principal se disponen las vesculas esfricas recin exocitadas. El aparato
de Golgi se puede dividir en tres regiones funcionales:
Regin Cis-Golgi: es la ms interna y prxima al retculo. De l recibe las
vesculas de transicin, que son sculos con protenas que han sido
sintetizadas en la membrana del retculo endoplasmtico rugoso (RER),
introducidas dentro de sus cavidades y transportadas por el lumen hasta
la parte ms externa del retculo. Estas vesculas de transicin son el
vehculo de dichas protenas que sern transportadas a la cara externa del
aparato de Golgi.
Regin medial: es una zona de transicin.
Regin Trans-Golgi: es la que se encuentra ms cerca de la membrana
plasmtica. De hecho, sus membranas, ambas unitarias, tienen una
composicin similar.
Las vesculas provenientes del retculo endoplsmico se fusionan con el
cis-Golgi, atravesando todos los dictiosomas hasta el trans-Golgi, donde
son empaquetadas y enviadas al lugar que les corresponda. Cada regin
contiene diferentes enzimas que modifican selectivamente las vesculas
segn donde estn destinadas.2 Sin embargo, an no se han logrado
determinar en detalle todas las funciones y estructuras del aparato de
Golgi.
Funciones generales
La clula sintetiza un gran nmero de diversas macromolculas necesarias
para la vida. El aparato de Golgi se encarga de la modificacin,
distribucin y envo de dichas macromolculas en la clula. Modifica
protenas y lpidos (grasas) que han sido sintetizados previamente tanto
en el retculo endoplasmtico rugoso como en el liso y los etiqueta para
enviarlos a donde corresponda, fuera o dentro de la clula. Las principales
funciones del aparato de Golgi vienen a ser las siguientes:
Modificacin de sustancias sintetizadas en el RER: En el aparato de Golgi
se transforman las sustancias procedentes del RER. Estas
transformaciones pueden ser agregaciones de restos de carbohidratos
para conseguir la estructura definitiva o para ser proteolizados y as
adquirir su conformacin activa. Por ejemplo, en el RER de las clulas
acinosas del pncreas se sintetiza la proinsulina que debido a las
transformaciones que sufre en el aparato de Golgi, adquirir la forma o
conformacin definitiva de la insulina. Las enzimas que se encuentran en
el interior de los dictiosomas son capaces de modificar las
macromolculas mediante glicosilacin (adicin de carbohidratos) y
fosforilacin (adicin de fosfatos). Para ello, el aparato de Golgi
transporta ciertas sustancias como nucletidos y azcares al interior del
orgnulo desde el citoplasma. Las protenas tambin son marcadas con
secuencias seal que determinan su destino final, como por ejemplo, la
manosa-6-fosfato que se aade a las protenas destinadas a los lisosomas.
Para llevar a cabo el proceso de fosforilacin el aparato de Golgi importa
molculas de ATP al interior del lumen,3 donde las kinasas catalizan la
reaccin. Algunas de las molculas fosforiladas en el aparato de Golgi son
las apolipoprotenas que dan lugar a las conocidas VLDL que se
encuentran en el plasma sanguneo. Parece ser que la fosforilacin de
estas molculas es necesaria para favorecer la secrecin de las mismas al
torrente sanguneo.
.4Secrecin celular: las sustancias atraviesan todos los sculos del aparato
de Golgi y cuando llegan a la cara trans del dictiosoma, en forma de
vesculas de secrecin, son transportadas a su destino fuera de la clula,
atravesando la membrana citoplasmtica por exocitosis. Un ejemplo de
esto son los proteoglicanos que conforman la matriz extracelular de los
animales. El aparato de Golgi es el orgnulo de mayor sntesis de
carbohidratos.5 Esto incluye la produccin de glicosaminoglicanos (GAGs),
largos polisacridos que son anclados a las protenas sintetizadas en el RE
para dar lugar a los proteoglicanos. De esto se encargarn las enzimas del
Golgi por medio de un residuo de xilosa. Otra forma de marcar una
protena puede ser por medio de la sulfatacin de una sulfotransferasa,
que gana una molcula de azufre de un donador denominado PAPs. Este
proceso tiene lugar en los GAGs de los proteoglicanos as como en los
ncleos de las protenas. Este nivel de sulfatacin es muy importante para
los proteoglicanos etiquetando funciones y dando una carga neta
negativa al proteoglicano.5
Produccin de membrana plasmtica: los grnulos de secrecin cuando se
unen a la membrana en la exocitosis pasan a formar parte de esta,
aumentando el volumen y la superficie de la clula.
Formacin de los lisosomas primarios.
Formacin del acrosoma de los espermios.
Vesculas de transporte[editar]
Las vesculas formadas en el retculo endoplasmtico liso forman,
unindose entre ellas, agregados tubulo-vesiculares, los cuales son
transportados hasta la regin cis del aparato de Golgi por protenas
motoras guiadas por microtbulos donde se fusionan con la membrana de
ste, vaciando su contenido en el interior del lumen. Una vez dentro, las
molculas son modificadas, marcadas y dirigidas hacia su destino final. El
aparato de Golgi tiende a ser mayor y ms numeroso en aquellas clulas
que sintetizan y secretan continuamente sustancias, como pueden ser los
linfocitos B y las clulas secretoras de anticuerpos.
Aquellas protenas destinadas a zonas alejadas del aparato de Golgi son
desplazadas hacia la regin trans, internndose en una compleja red de
membranas y vesculas asociadas denominadas regin trans-Golgi.6 Esta
regin es donde muchas protenas son marcadas y enviadas hacia sus
correspondientes destinos por medio de alguno de estos 3 tipos
diferentes de vesculas, segn el marcador que presenten
MODELOS DE TRANSPORTE
Los mecanismos de transporte que utilizan las protenas para trasladarse
a travs del aparato de Golgi no estn muy claros an, por lo que existen
diversas hiptesis para explicar dicho desplazamiento. Actualmente,
existen dos modelos predominantes que no son excluyentes entre s,
hasta el punto de ser referidos a veces como el modelo combinado.5
*Modelo de maduracin de las cisternas: las cisternas del aparato de
Golgi llevan cabo un movimiento unidireccional desde la regin cis, donde
se forman, hasta la regin trans, donde son destruidas. Las vesculas del
retculo endoplasmtico se fusionan con los dictiosomas de la regin cis
para dar lugar a nuevas cisternas, lo que podra generar el movimiento de
las cisternas a travs del aparato de Golgi a medida que se van formando
nuevas cisternas en la regin cis. Este modelo se apoya en el hecho de
que se han observado al microscopio estructuras mayores que las
vesculas de transporte, tales como las fibras de colgeno, desplazndose
a travs del aparato de Golgi.5 Inicialmente, esta hiptesis tuvo una gran
acogida y fue la ms aceptada hasta los aos 80. Los ltimos estudios
realizados al respecto por la Universidad de Tokio y la Universidad de
Chicago con tecnologa ms avanzada han permitido observar con mayor
detalle los compartimentos y el proceso de maduracin del aparato de
Golgi.7 Adems, existen evidencias de movimientos retrgrados (en
direccin cis) de cierto tipo de vesculas (COP1), que transportan
protenas del retculo endoplasmtico mediante el reconocimiento de
pptidos seales.8
*Modelo del transporte vesicular: el transporte vesicular asume que el
aparato de Golgi es un orgnulo muy estable y esttico, dividido en
compartimentos que se disponen en direccin cis trans. Las vesculas
son las encargadas de transportar el material entre el retculo
endoplasmtico y el aparato de Golgi y entre los diferentes
compartimentos de este.9 Las evidencias experimentales que apoyan esta
hiptesis se basan en la gran abundancia de vesculas pequeas
(conocidas tcnicamente como vesculas lanzadera) localizadas en las
proximidades del aparato de Golgi. La direccionalidad vendra dada por
las protenas trasportadas en el interior de las vesculas, cuyo destino
determinara el movimiento de avance o de retroceso a travs del aparato
de Golgi, aunque tambin podra suceder que la direccionalidad no fuera
necesaria y el destino de las protenas viniera ya determinado desde el
retculo endoplasmtico. Al margen de esto, es probable que el transporte
de vesculas se encuentre asociado al citoesqueleto por medio de
filamentos de actina, encargados de asegurar la fusin de las vesculas con
sus correspondientes compartimentos.5
TRANSPORTE VESICULAR
La incorporacin a la clula de molculas de gran tamao (enzimas, cidos
nucleicos, etc) se realiza por un mecanismo de vesiculacin denominado
endocitosis. Estas macromolculas tambin pueden ser segregadas cara el
exterior por un proceso inverso denominado exocitosis, son transportes
que conllevan gastos de energa.
Endocitosis. Implica un englobamiento del material a transportar en una
invaginacin de la membrana plasmtica. Se va a formar una vescula
cerrada que se libera al interior de la cula y que queda separada del
citosol por una membrana. Si las partculas que se incorporan por
vesiculacin son slidas y de gran tamao, se habla de fagocitosis, si son
pequeas gotas de lquido extracelular o macromolculas se habla de
pinocitosis.
En el proceso de endocitosis los ligandos (material que va a ser
fagocitado) se van acoplar a los receptores de la membrana plasmtica
llamados tambin receptores de carga. Estos receptores son capturados
por las adaptinas que se van a unir a las molculas de Clatrina formando
vesculas recubiertas por clatrina. La protena dinamina se ensambla en el
cuello de las vesculas que se estn formando y la separan. Despus se
eliminan las protenas de cubierta y la membrana plasmtica de la
vescula se puede fusionar con su membrana diana.
Las vesculas se fusionan con otros similares formando endosomas.
Primero van a ser endosomas tempranos, situados en la periferia de la
clula y despues van a ser endosomas tardos, en los que se desacopla el
receptor y el ligando y estos endosomas tardos se van a situar cerca del
ncleo o del aparato de Golgi.
Todo el proceso de endocitosis consume energa. Las funciones mediadas
por endocitose pueden ser varias:
1- funcin de digestin celular: los endosomas se van a fusionar con
lisosomas formando endolisosomas. En los endolisosomas se degradan las
molculas incorporadas y sus productos pasan a travs de la membrana
endolisosomal cara el citosol donde pueden ser utilizados.
2- Reciclaje de la membrana plasmtica.
3- Almacenamiento de sustancia de reserva (ej: los lpidos).
4- Transporte vesicular: llamado tambin transcitosis. Es el transporte de
sustancias de un lugar la otro de la clula. En este transporte, las vesculas
de transporte presentan marcadores vesiculares que parece que son unas
protenas transmembrana denominadas Snare. Las vesculas que emergen
contienen protenas marcadoras llamadas snare vesiculares o V-snare que
se van a unir a la snare diana o T-snare de la membrana diana. Tras lo
acoplamento de una vescula con la membrana diana, un complejo de
protenas de fusin de la membrana ensmblase en el punto de
acoplamento y catalizan la fusin de la vescula con la membrana diana.
Fagocitosis.
Este proceso es similar en todas las clulas fagocitarias (neutrfilos o
macrfogos). Para captar partculas de grano tamao (bacterias) las
clulas emiten proyecciones. Estas rodean las partculas cerrndose sobre
ellas formando una vacuola de endocitosis que se separa de la membrana
plasmtica y se introduce en el citoplasma.
Las clulas fagocticas poseen tanto receptores para los anticuerpos que
se unieron a los antigenos de la partcula fagocitada cmo para el
componente C3 del complemento que tambin se une a los antgenos.
Mediante estos receptores, los fagocitos se van a unir la partcula que van
fagocitar. En la degradacin del material fagocitado intervienen los
lisosomas que contienen enzimas hidrolticas. Estos lisosomas se unen a
las vacuolas de fagocitosis formando una vacuola digestiva llamada
fagolisosoma.
2- Exocitosis.
Proceso inverso a la endocitosis. Es utilizado para que la clula vierta al
exterior diversas sustancias (enzimas o hormonas).
El proceso ms representativo es la secrecin celular. Hay dos tipos de
secrecin:
1- Secrecin constitutiva: es realizada por todas las clulas y de modo
continuo mediante vesculas que proceden del complejo de Golgi y que se
fusionan con la membrana plasmtica, corresponde a sustancias que van
destinadas al medio extracelular (ej: proteogluconas y protenas de la
matriz extracelular, o tambin protenas de la membrana que se van a
quedar en la superficie celular). Tambin se incluye en este tipo de
secrecin la regeneracin de la bicapa lipdica y del glucocalix.
2- Secrecin regulada: se produce solo en algunas clulas llamadas clulas
secretoras y solo cuando la clula es estimulada por una seal
extracelular, generalmente un mensajero qumico, que se une a los
receptores de la superficie celular e induce cambios intracelulares como
un aumento en la concentracion de ion calcio y que hace que las
protenas se agreguen. Estas protenas agregadas son reconocidas,
empaquetadas en vesculas y liberadas.
Cuando una vescula de secrecin se fusiona con la membrana plasmtica
y descarga su contenido por exocitose, su membrana se vuelve parte de la
membrana plasmtica. Aun que esto debera aumentar la superficie de la
membrana plasmtica, slo es transitoriamente porque los componentes
de la membrana son eliminados en otras regiones por endocitosis,
habiendo asi una renovacin contnua de los lpidos y protenas de
membrana.
LISOSOMAS
Los lisosomas tienen una estructura muy sencilla, "semejantes" a
vacuolas, rodeados solamente por una membrana, contienen gran
cantidad de enzimas digestivas que degradan todas las molculas
inservibles para la clula.
Funcionan como "estmagos" de la clula y adems de digerir cualquier
sustancia que ingrese del exterior, vacuolas digestivas (figura, nmeros 4
y 5), ingieren restos celulares viejos para digerirlos tambin (nmero 3),
llamados entonces vacuolas autofgicas.
Llamados "bolsas suicidas" porque si se rompiera su membrana, las
enzimas encerradas en su interior, terminaran por destruir a toda la
clula.
Los lisosomas se forman a partir del Retculo endoplsmico rugoso
(nmero 1)y posteriormente las enzimas son empaquetadas por el
Complejo de Golgi (nmero 2).
Son vesculas englobadas por una membrana que se forman en la aparato
de Golgi y que contienen un gran nmero de enzimas digestivas
(hidrolticas y proteolticas) capaces de romper una gran variedad de
molculas. La carencia de algunas de estas enzimas puede ocasionar
enfermedades metablicas como la enfermedad de Tay-Sachs.
Las enzimas proteolticas funcionan mejor a pH cido y, para conseguirlo
la membrana del lisosoma contiene una bomba de protones que
introduce H+ en la vescula. Como consecuencia de esto, el lisosoma tiene
un pH inferior a 5.0.
Las enzimas lisosomales son capaces de digerir bacterias y otras
sustancias que entran en la clula por fagocitosis, u otros procesos de
endocitosis. Eventualmente, los productos de la digestin son tan
pequeos que pueden pasar la membrana del lisosoma volviendo al
citosol donde son recicladas Los lisosomas utilizan sus enzimas para
reciclar los diferentes orgnulos de la clula, englobndolos, digirindoles
y liberando sus componentes en el citosol. De esta forma los orgnulos de
la clula se estn continuamente reponiendo. El proceso de digestin de
los organuelos se llama autofagia. Por ejemplo, las clulas hepticas se
reconstituyen por completo una vez cada dos semanas. Otra funcin de
los lisosomas es la digestin de detritus extracelulares en heridas y
quemaduras, preparando y limpiando el terreno para la reparacin del
tejido. Se debe tener en cuenta que las protenas que integran la
membrana del lisosoma son altamente glicosiladas, para evitar ser
degradadas por las proteasas del lumen.
LA VIA ENDOCITICA
La clula es un sistema abierto que requiere del intercambio de materia y
energa con el exterior para mantener su viabilidad. Aunque las protenas
transportadoras situadas en la membrana plasmtica median el
movimiento de iones y pequeas molculas a travs de la bicapa lipdica,
la clula internaliza grandes macromolculas solubles del medio
extracelular a travs del proceso de endocitosis que se desarrolla a lo
largo de la Va endoctica
*Endocitosis
Endocitosis del griego endo, dentro, cito clula, y sis accin, (literalmente
accin de introducir en la clula) es el proceso pues por el que la clula
toma material desde el exterior celular. En la endocitosis el material que
se va a introducir es rodeado por una porcin de la membrana plasmtica
que luego se invagina para generar por pellizco y pinzamiento regulado
una porcin de la membrana plasmtica que termina generando una
vescula endoctica que contiene el material internalizado (llamado
generalmente cargo). Esta vescula endoctica ser procesada
intracelularmente en una serie compartimentos endocticos intracelulares
de la va endosomal donde el cargo contenido de la misma ser
procesado.
*Tipos de endocitosis
Por la manera en el que las vesculas endocticas son formadas, el tipo de
material que es internalizado, as como el tamao de las mismas, se
puede distinguir varios tipos de endocitosis:
*Fagocitosis
Fagocitosis, del griego fago, comer, cito clula sis, accin (literalmente
accin celular de comer), es la internalizacin por la clula de grandes
partculas, usualmente grandes agregados de macromolculas (e.g.
complejos antgenos-anticuerpo) o de microorganismos (bacterias) del
entorno extracelular. En los organismos multicelulares, la fagocitosis es
llevada a cabo por clulas especializadas como los macrfagos. Este tipo
clulas son glbulos blancos, leucocitos, de la sangre que reconocen,
engullen y destruyen bacterias; eliminan tambin desechos celulares o
incluso clulas intactas (e.g. eritrocitos o glbulos rojos viejos). Estas
clulas fagocitarias reciben por ello el nombre de fagotitos profesionales,
y constituyen una parte esencial del sistema inmunitario innato del
organismo, un sistema de defensa que combate a los microorganismos
que infectan e invaden el organismo.
El proceso de fagocitosis se pone en marcha cuando una bacteria por
ejemplo interacciona con receptores de la membrana plasmtica del
macrfago lo que dispara la extensin (en un movimiento de dependiente
de la polimerizacin de filamentos de actina) de protuberancias en la
superficie celular llamadas pseudopodos (del griego, pseudo falso y podo
pie, literalmente falsos pies). Los pseudpodos acaban rodeando a la
bacteria que terminan siendo engullida y encapsulada en una vescula
endoctica intracelular especializada de gran tamao (>0.25 micras de
dimetro) llamada fagosoma. El fagosoma se fusiona entonces con un
lisosoma primario el sufre un proceso de maduracin para producir a un
lisosoma maduro llamado ahora fagolisosoma en la cual se digerir
(destruir) la bacteria que ha sido capturada e internalizada. En
organismos unicelulares como las amebas la fagocitosis es empleada para
capturar partculas alimenticias, como bacterias u otros protozoos.
*Pinocitosis
Pinocitosis, del griego pino, beber, cito clula, y sis, accin (literalmente la
accin celular de beber) es la internalizacin de gotas de fluido
extracelular, es la ms comn en las clulas eucariotas. Pinocitosis se
refiere a la formacin constitutiva de pequeas vesculas que llevan fluido
extracelular con macromolculas que pueden estar no estar
especficamente unidas a receptores presentes en la membrana
plasmtica. El termino pinocitosis est cada vez ms en desuso y estos
procesos son referidos generalmente con el termino general de
endocitosis
Você também pode gostar
- Plan de Trabajo SaludDocumento10 páginasPlan de Trabajo Saludenzosp18Ainda não há avaliações
- Oficio EnzoDocumento1 páginaOficio Enzoenzosp18Ainda não há avaliações
- Lambayeque PEES 2016-2018Documento20 páginasLambayeque PEES 2016-2018enzosp180% (1)
- Procesadora Peru SDocumento3 páginasProcesadora Peru Senzosp18Ainda não há avaliações
- MampostDocumento1 páginaMampostMisaelPGAinda não há avaliações
- Roundcube Webmail - Propuesta de Agenda Reunion 05-08-2016Documento1 páginaRoundcube Webmail - Propuesta de Agenda Reunion 05-08-2016enzosp18Ainda não há avaliações
- Rad 5 F 810Documento177 páginasRad 5 F 810enzosp18Ainda não há avaliações
- Análisis de Ratios de Gestion - Aceros Arequipa S.A.Documento9 páginasAnálisis de Ratios de Gestion - Aceros Arequipa S.A.enzosp18Ainda não há avaliações
- Es Rentable Tu Negocio Gustavo SamaniegoDocumento32 páginasEs Rentable Tu Negocio Gustavo Samaniegoenzosp18Ainda não há avaliações
- Evlución de ProcesosDocumento1 páginaEvlución de Procesosenzosp18Ainda não há avaliações
- Calorimetria PDFDocumento4 páginasCalorimetria PDFMaritZa JulcaAinda não há avaliações
- Matris Biz 1 PDFDocumento12 páginasMatris Biz 1 PDFenzosp18Ainda não há avaliações
- Sesión 01 - Finanzas InternacionalesDocumento32 páginasSesión 01 - Finanzas Internacionalesenzosp180% (1)
- Monografia REPRODUCCIÓN ASEXUAL DIRECTA E INDIRECTADocumento29 páginasMonografia REPRODUCCIÓN ASEXUAL DIRECTA E INDIRECTAEder Aquino GalarzaAinda não há avaliações
- Silabo 1647828770485Documento6 páginasSilabo 1647828770485Héctor reyes Garcia Reyes GarciaAinda não há avaliações
- NematodosDocumento17 páginasNematodosNatalia Gonzalez PabonAinda não há avaliações
- Mangure Pisangio 3Documento9 páginasMangure Pisangio 3olgerAinda não há avaliações
- PermeabilidadDocumento21 páginasPermeabilidadeleidaAinda não há avaliações
- Biologia Tarea 4Documento13 páginasBiologia Tarea 4JEOVANNYAinda não há avaliações
- Oiv Viti 564b 2019 EsDocumento19 páginasOiv Viti 564b 2019 EsJesúsAinda não há avaliações
- Neuronas piramidales: funciones clave en el cerebroDocumento4 páginasNeuronas piramidales: funciones clave en el cerebromayAinda não há avaliações
- Tinción Gram: diferenciar bacterias enDocumento6 páginasTinción Gram: diferenciar bacterias enLorena Medaly Bendezu PaucarAinda não há avaliações
- Reino ProtistaDocumento6 páginasReino ProtistaNorma Luz FajardoAinda não há avaliações
- Resveratrol PDFDocumento70 páginasResveratrol PDFMonicaAinda não há avaliações
- López Araceli Enfermedades ViralesDocumento7 páginasLópez Araceli Enfermedades ViralesAra LopezAinda não há avaliações
- V. Ev 1 CatálogoDocumento28 páginasV. Ev 1 Catálogojoselyne luqueAinda não há avaliações
- La Fotosíntesis de La IngenieríaDocumento2 páginasLa Fotosíntesis de La IngenieríaMario Arriola VertizAinda não há avaliações
- Copia de Martínez Hernáez, A. (2008) - La Enfermedad Vista Por Los Antropólogos. en Antropología Médica. Teoría Sobre La Cultura, El Poder y La EnfermedadDocumento11 páginasCopia de Martínez Hernáez, A. (2008) - La Enfermedad Vista Por Los Antropólogos. en Antropología Médica. Teoría Sobre La Cultura, El Poder y La EnfermedadCristian Nicolás HerreraAinda não há avaliações
- Apoped FisioDocumento211 páginasApoped FisioDiana SalazarAinda não há avaliações
- Crecimiento y DesarrolloDocumento29 páginasCrecimiento y DesarrolloConyMaldonadoAinda não há avaliações
- Tema 1 Introduccion Al Metabolismo y Señalización Celular 2018Documento65 páginasTema 1 Introduccion Al Metabolismo y Señalización Celular 2018uniautonoma53Ainda não há avaliações
- Inmunologia CompletoDocumento59 páginasInmunologia CompletoSebastián AgudeloAinda não há avaliações
- Chi CuadradoDocumento74 páginasChi CuadradoYris RosarioAinda não há avaliações
- Actualizacion en El Diagnóstico Y Tratamiento de Las Meningoencefalitis Asépticas Del PerroDocumento14 páginasActualizacion en El Diagnóstico Y Tratamiento de Las Meningoencefalitis Asépticas Del PerroSamanthaRoqueAinda não há avaliações
- CriptosporidiosisDocumento32 páginasCriptosporidiosisDaira CantuAinda não há avaliações
- Norma Nacional Reglas Protocolos y Procedmientos para La DetDocumento73 páginasNorma Nacional Reglas Protocolos y Procedmientos para La DetOlivert Nelson Rios JustinianoAinda não há avaliações
- Actividad-Taller N°1 Historia MicrobiologiaDocumento15 páginasActividad-Taller N°1 Historia MicrobiologiagletopeAinda não há avaliações
- Ecología y Conservación de Bosques Neotropicales (Manuel R. Guariguata, Gustavo H. Kattan)Documento347 páginasEcología y Conservación de Bosques Neotropicales (Manuel R. Guariguata, Gustavo H. Kattan)B AleSis Morales100% (1)
- Espermatogénesis: proceso de formación de espermatozoidesDocumento5 páginasEspermatogénesis: proceso de formación de espermatozoidesKarla Quiñones RufinoAinda não há avaliações
- Monografia BioseguridadDocumento23 páginasMonografia BioseguridadMadHe ValdiiviiaAinda não há avaliações
- Silabo de Patologia GeneralDocumento18 páginasSilabo de Patologia GeneralEdwin Morales CarboneAinda não há avaliações
- Estructura nuclearDocumento5 páginasEstructura nuclearAndres Felipe Prieto AlarconAinda não há avaliações
- Hist Med PrehistDocumento2 páginasHist Med PrehistDomna VelizAinda não há avaliações