Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Bio 3100 Biochemistry Palomino Exams BlankDocumento14 páginasBio 3100 Biochemistry Palomino Exams Blankteamaqua360100% (1)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- C.5 Encopresis 0072012Documento14 páginasC.5 Encopresis 0072012Indah Afriani Nasution100% (1)
- C.5 Encopresis 0072012Documento14 páginasC.5 Encopresis 0072012Indah Afriani Nasution100% (1)
- Underwater Seal Chest DrainageDocumento22 páginasUnderwater Seal Chest DrainageRegina SeptianiAinda não há avaliações
- JOPH2015 509456editDocumento7 páginasJOPH2015 509456editRegina SeptianiAinda não há avaliações
- Practice: Bilateral Peritonsillar Abscesses Complicating Acute TonsillitisDocumento4 páginasPractice: Bilateral Peritonsillar Abscesses Complicating Acute TonsillitisRegina SeptianiAinda não há avaliações
- Review Article: Uveitis in The Aging Eye: Incidence, Patterns, and Differential DiagnosisDocumento8 páginasReview Article: Uveitis in The Aging Eye: Incidence, Patterns, and Differential DiagnosisRegina SeptianiAinda não há avaliações
- The Factor Structure For The Positive and Negative Syndrome ScaleDocumento2 páginasThe Factor Structure For The Positive and Negative Syndrome ScaleRegina SeptianiAinda não há avaliações
- Case Report: Plasenta PreviaDocumento2 páginasCase Report: Plasenta PreviaRegina SeptianiAinda não há avaliações
- Ni Hms 597871Documento20 páginasNi Hms 597871Regina SeptianiAinda não há avaliações
- Kuliah Audiologi-1Documento36 páginasKuliah Audiologi-1Regina SeptianiAinda não há avaliações
- The Factor Structure For The Positive and Negative Syndrome ScaleDocumento2 páginasThe Factor Structure For The Positive and Negative Syndrome ScaleRegina SeptianiAinda não há avaliações
- Denah IKMDocumento1 páginaDenah IKMRegina SeptianiAinda não há avaliações
- The Factor Structure For The Positive and Negative Syndrome ScaleDocumento2 páginasThe Factor Structure For The Positive and Negative Syndrome ScaleRegina SeptianiAinda não há avaliações
- Underwater Seal Chest DrainageDocumento22 páginasUnderwater Seal Chest DrainageRegina SeptianiAinda não há avaliações
- Div Styletext-Align Justify Biochemistry of Magnesium-DivDocumento22 páginasDiv Styletext-Align Justify Biochemistry of Magnesium-DivRegina SeptianiAinda não há avaliações
- Death After Head Injury: The 13 Year Outcome of A Case Control StudyDocumento6 páginasDeath After Head Injury: The 13 Year Outcome of A Case Control StudyRegina SeptianiAinda não há avaliações
- Enzim KlinikDocumento20 páginasEnzim KlinikRegina SeptianiAinda não há avaliações
- Enzyme - 2: Aspect Molecular of EnzymeDocumento14 páginasEnzyme - 2: Aspect Molecular of EnzymeRegina SeptianiAinda não há avaliações
- Jurnal Reading: Efek ACE Inhibitor (Zofenopril) Terhadap Mortalitas Dan Morbiditas Setelah Infark Miokard AnteriorDocumento1 páginaJurnal Reading: Efek ACE Inhibitor (Zofenopril) Terhadap Mortalitas Dan Morbiditas Setelah Infark Miokard AnteriorRegina SeptianiAinda não há avaliações
- Nutrients 05 03910Documento10 páginasNutrients 05 03910Regina SeptianiAinda não há avaliações
- Enzyme - 2: Aspect Molecular of EnzymeDocumento14 páginasEnzyme - 2: Aspect Molecular of EnzymeRegina SeptianiAinda não há avaliações
- Nutrients 05 03910Documento10 páginasNutrients 05 03910Regina SeptianiAinda não há avaliações
- Underwater Seal Chest DrainageDocumento22 páginasUnderwater Seal Chest DrainageRegina SeptianiAinda não há avaliações
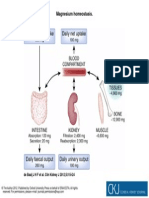
- Clin Kidney J 2012 Feb 5 (Suppl 1) I15-24, Fig. 1Documento1 páginaClin Kidney J 2012 Feb 5 (Suppl 1) I15-24, Fig. 1Regina SeptianiAinda não há avaliações
- SoeriptoDocumento8 páginasSoeriptoRegina SeptianiAinda não há avaliações
- PolymorphDocumento17 páginasPolymorphRegina SeptianiAinda não há avaliações
- 418 Jadwal Pendaftaran Beasiswa Program Sarjana 1310031017Documento1 página418 Jadwal Pendaftaran Beasiswa Program Sarjana 1310031017Regina SeptianiAinda não há avaliações
- Emergency THTDocumento37 páginasEmergency THTIndah D. RahmahAinda não há avaliações
- NIH Public Access: Author ManuscriptDocumento14 páginasNIH Public Access: Author ManuscriptRegina SeptianiAinda não há avaliações
- Cell Physiology Week 2 Revised Tabrez Specialized Organels-Cytoplasm 1445 HDocumento40 páginasCell Physiology Week 2 Revised Tabrez Specialized Organels-Cytoplasm 1445 H6258c7pxkjAinda não há avaliações
- BIO504-Practical Exam Notes VUDocumento34 páginasBIO504-Practical Exam Notes VUSagheer AhmedAinda não há avaliações
- Water Soluble VitaminsDocumento36 páginasWater Soluble VitaminsMehak ElahiAinda não há avaliações
- HL Paper 2: (8 Max) If Definition Is MissingDocumento55 páginasHL Paper 2: (8 Max) If Definition Is MissingJane ChangAinda não há avaliações
- Lipids and LipoproteinsDocumento5 páginasLipids and LipoproteinsMica BernardoAinda não há avaliações
- Nephrotic SyndromeDocumento5 páginasNephrotic SyndromeMahmoud SalmanAinda não há avaliações
- Cell 101029Documento196 páginasCell 101029PrasathAinda não há avaliações
- DIALAB Chemistry 2016 PDFDocumento2 páginasDIALAB Chemistry 2016 PDFKhaled AlkhawaldehAinda não há avaliações
- Computeraideddrugdesign 130701054152 Phpapp01Documento61 páginasComputeraideddrugdesign 130701054152 Phpapp01Silverstar AzarAinda não há avaliações
- CatSper Calcium Channels: 20 Years OnDocumento16 páginasCatSper Calcium Channels: 20 Years OnBiblioteca Campus JuriquillaAinda não há avaliações
- Chapter 8 Cell Reproduction Study GuideDocumento2 páginasChapter 8 Cell Reproduction Study GuidemarigigAinda não há avaliações
- Buletin de Analize Medicale NR 1827118: Nume Pacient Prenume Pacient Sex Data NasteriiDocumento3 páginasBuletin de Analize Medicale NR 1827118: Nume Pacient Prenume Pacient Sex Data NasteriiIulian CezarAinda não há avaliações
- Cell Model Rubric: Material GuidelinesDocumento2 páginasCell Model Rubric: Material GuidelinesHazel Jane MalicdemAinda não há avaliações
- Chapter 01. Introduction To Biopharmaceutics and Its Role in Drug DevelopmentDocumento25 páginasChapter 01. Introduction To Biopharmaceutics and Its Role in Drug DevelopmentSava19880% (1)
- DNA Replication and Protein SynthesisDocumento34 páginasDNA Replication and Protein SynthesisJessa Mae Infante PadillaAinda não há avaliações
- Glycolysis, Kreb Cycle Electron TransportDocumento19 páginasGlycolysis, Kreb Cycle Electron TransportShahidatul4297100% (2)
- 10.1. Living Organisms and Life Processes - Centre of Excellence Academy PDFDocumento5 páginas10.1. Living Organisms and Life Processes - Centre of Excellence Academy PDFFrancis JrAinda não há avaliações
- CRISPRDocumento6 páginasCRISPRvijayjeoAinda não há avaliações
- Sephadex LH-20: Gel FiltrationDocumento8 páginasSephadex LH-20: Gel FiltrationUmi Darozah ZainAinda não há avaliações
- KERAGAMAN GENETIK GURITA Octopus Cyanea (LINNAEUS 1758) PAPUADocumento9 páginasKERAGAMAN GENETIK GURITA Octopus Cyanea (LINNAEUS 1758) PAPUAJen SajaAinda não há avaliações
- Assay Development and High-ThroughputDocumento12 páginasAssay Development and High-ThroughputJuan AspilcuetaAinda não há avaliações
- Chapter 2 The Cytoplasm ReviewerDocumento13 páginasChapter 2 The Cytoplasm ReviewerKyleBernalÜAinda não há avaliações
- CAPE Biology 2010 U2 P2 MSDocumento15 páginasCAPE Biology 2010 U2 P2 MSYagna Lall100% (1)
- Chapman 1978Documento5 páginasChapman 1978aneupane465Ainda não há avaliações
- Nameer Taha Al Hayali - 3622103731Documento2 páginasNameer Taha Al Hayali - 3622103731Nameer omarAinda não há avaliações
- Bio-Data: Degree Subject Area Period of Study University / Institute Subjects From ToDocumento7 páginasBio-Data: Degree Subject Area Period of Study University / Institute Subjects From ToampharmAinda não há avaliações
- Tribhuvan University Institute of Science and Technology: Course of Study For Four Year B. Sc. MicrobiologyDocumento23 páginasTribhuvan University Institute of Science and Technology: Course of Study For Four Year B. Sc. MicrobiologyRanjit Kumar ShahAinda não há avaliações
- L2 Genome OrganizationDocumento18 páginasL2 Genome OrganizationGaayithri RAinda não há avaliações
- Lehninger Principles of Biochemistry, Fourth Edition - David L. Nelson, Michael M. Cox (0354-0377) (13-24)Documento12 páginasLehninger Principles of Biochemistry, Fourth Edition - David L. Nelson, Michael M. Cox (0354-0377) (13-24)Nur AuliaAinda não há avaliações