Você também pode gostar
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Belt Failure AnalysisDocumento8 páginasBelt Failure AnalysisDavid ChandlerAinda não há avaliações
- Intro To Dental AnatomyDocumento15 páginasIntro To Dental AnatomydentalA5455Ainda não há avaliações
- Advanced Tooth Morphology, Tooth Identification and Enamel COH Senior StudentsDocumento10 páginasAdvanced Tooth Morphology, Tooth Identification and Enamel COH Senior StudentsshuankayAinda não há avaliações
- Speciation Natural Processes, Genetics and BiodiversityDocumento276 páginasSpeciation Natural Processes, Genetics and BiodiversityCristian BarrosAinda não há avaliações
- Effective Exercises For A Short FrenumDocumento2 páginasEffective Exercises For A Short FrenumlblessingerAinda não há avaliações
- Experimental Gingivitis Study Links Oral Bacteria to InflammationDocumento11 páginasExperimental Gingivitis Study Links Oral Bacteria to InflammationCassia Martins PereiraAinda não há avaliações
- Esthetic Dentistry / Orthodontic Courses by Indian Dental AcademyDocumento256 páginasEsthetic Dentistry / Orthodontic Courses by Indian Dental Academyindian dental academyAinda não há avaliações
- Oral Histology Quiz - Scientific Term (AmCoFam)Documento22 páginasOral Histology Quiz - Scientific Term (AmCoFam)AmericanCornerFamily100% (5)
- QDT 2001 PDFDocumento239 páginasQDT 2001 PDFAlex BurdeAinda não há avaliações
- Eutheria (Placental Mammals) : J David ArchibaldDocumento4 páginasEutheria (Placental Mammals) : J David Archibaldmanoj_rkl_07Ainda não há avaliações
- Certificate of Foreign Status of Beneficial Owner For United States Tax Withholding and Reporting (Individuals)Documento1 páginaCertificate of Foreign Status of Beneficial Owner For United States Tax Withholding and Reporting (Individuals)Cristian BarrosAinda não há avaliações
- Structural Characteristics of The Hair of MammalsDocumento29 páginasStructural Characteristics of The Hair of MammalsCristian BarrosAinda não há avaliações
- A Practical Guide To The Application of The IUCN Red List of Ecosystems CriteriaDocumento9 páginasA Practical Guide To The Application of The IUCN Red List of Ecosystems CriteriaCristian BarrosAinda não há avaliações
- Bhosques SecosDocumento26 páginasBhosques SecosRuslan SokolovAinda não há avaliações
- Bhosques SecosDocumento26 páginasBhosques SecosRuslan SokolovAinda não há avaliações
- The Use of Camera Traps and Mammal TrackDocumento16 páginasThe Use of Camera Traps and Mammal TrackCristian BarrosAinda não há avaliações
- Maximum Estimated Bite Force, Skull Morphology, and Primary Prey Size in North American CarnivoresDocumento179 páginasMaximum Estimated Bite Force, Skull Morphology, and Primary Prey Size in North American CarnivoresCristian BarrosAinda não há avaliações
- Conservation Value of Forest Fragments For Medium-Sized Carnivores in A Silvopastoral System in ColombiaDocumento5 páginasConservation Value of Forest Fragments For Medium-Sized Carnivores in A Silvopastoral System in ColombiaCristian BarrosAinda não há avaliações
- Intraspecific Killing of A Male OcelotDocumento3 páginasIntraspecific Killing of A Male OcelotCristian BarrosAinda não há avaliações
- Molecules Consolidate The Placental Mammal TreeDocumento9 páginasMolecules Consolidate The Placental Mammal TreeEduardo Antonio Molinari NovoaAinda não há avaliações
- Cámara Trampa BushellDocumento128 páginasCámara Trampa BushellCristian BarrosAinda não há avaliações
- Abreu Et Al 2008 Feeding Habits of Ocelot (Leopardus Pardalis) in Southern BrazilDocumento5 páginasAbreu Et Al 2008 Feeding Habits of Ocelot (Leopardus Pardalis) in Southern BrazilRodrigo MoroAinda não há avaliações
- Fototrampeo Del OceloteDocumento150 páginasFototrampeo Del OceloteCristian BarrosAinda não há avaliações
- Tooth Numbering SystemsDocumento13 páginasTooth Numbering Systemsdr parveen bathlaAinda não há avaliações
- The clothes will be dry after hanging outsideDocumento61 páginasThe clothes will be dry after hanging outsideWindel Beth Quimat ZafraAinda não há avaliações
- Teeth Feature Extraction and Matching For HumanDocumento6 páginasTeeth Feature Extraction and Matching For HumanBudi PurnomoAinda não há avaliações
- ICDAS Clinical ChartDocumento2 páginasICDAS Clinical Chartpiece_of_mindzz1969Ainda não há avaliações
- Kishen Hydormechanics in Dentine Role of Dentinal Tubules and Hydrostatic Pressure On Mechanical Stress-Strain DistributionDocumento11 páginasKishen Hydormechanics in Dentine Role of Dentinal Tubules and Hydrostatic Pressure On Mechanical Stress-Strain DistributionОлександр БайдоAinda não há avaliações
- Post-Traumatic Apexogenesis of Immature IncisorsDocumento4 páginasPost-Traumatic Apexogenesis of Immature IncisorsDiandra Amalia SuyudiAinda não há avaliações
- A panoramic view of tooth wear in medieval EuropeDocumento16 páginasA panoramic view of tooth wear in medieval EuropeLucas RossiAinda não há avaliações
- Zebra XX, Part 2 PDFDocumento3 páginasZebra XX, Part 2 PDFRamona MateiAinda não há avaliações
- Kings Way Primary School P4 English Holiday Work Week 3 Complete The Table Correctly. Masculine FeminineDocumento21 páginasKings Way Primary School P4 English Holiday Work Week 3 Complete The Table Correctly. Masculine FeminineMonydit santinoAinda não há avaliações
- Preventive Orthodontics / Orthodontic Courses by Indian Dental AcademyDocumento15 páginasPreventive Orthodontics / Orthodontic Courses by Indian Dental Academyindian dental academyAinda não há avaliações
- Stainless Steel Crown PostDocumento1 páginaStainless Steel Crown PostAgung AdhaAinda não há avaliações
- Tooth Development and Supporting TissuesDocumento68 páginasTooth Development and Supporting TissuesSaud IqbalAinda não há avaliações
- Envi285 Pset 6Documento5 páginasEnvi285 Pset 6JUSTINE ALEXIS CABONCEAinda não há avaliações
- BiologyDocumento39 páginasBiologyNil KamalAinda não há avaliações
- Principles of Tooth PreparationDocumento3 páginasPrinciples of Tooth PreparationMarian TavassoliAinda não há avaliações
- Amalgam Class ViDocumento14 páginasAmalgam Class ViJamaica Faith RamonAinda não há avaliações
- Eruption and Shedding of Teeth - 2014Documento70 páginasEruption and Shedding of Teeth - 2014leeminhoangrybirdAinda não há avaliações
- Grade 5 1st Term 2019-2020Documento85 páginasGrade 5 1st Term 2019-2020Hicham HamdyAinda não há avaliações
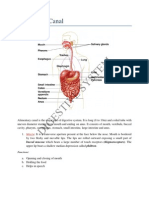
- Digestive System (Alimentary Canal + Digestive Glands + Digestive Process)Documento26 páginasDigestive System (Alimentary Canal + Digestive Glands + Digestive Process)Shuvanjan Dahal100% (1)
- BIOLOGY SS 1 WEEK 5 .PPTX EDocumento18 páginasBIOLOGY SS 1 WEEK 5 .PPTX EOfficial PageAinda não há avaliações
- Digital Dental Photography. Part 8: Intra-Oral Set-Ups: I. AhmadDocumento7 páginasDigital Dental Photography. Part 8: Intra-Oral Set-Ups: I. AhmadbkprosthoAinda não há avaliações
- Science Around Us Book 3 PDFDocumento94 páginasScience Around Us Book 3 PDFAbina VancootenAinda não há avaliações