Você também pode gostar
- Expo EnvasesDocumento12 páginasExpo EnvasesCordova FernandoAinda não há avaliações
- Trigo y DerivadosDocumento49 páginasTrigo y DerivadosCordova FernandoAinda não há avaliações
- PANIFICACIONDocumento10 páginasPANIFICACIONCordova FernandoAinda não há avaliações
- Expo CelmiraDocumento20 páginasExpo CelmiraCordova FernandoAinda não há avaliações
- Nuggets de PescadoDocumento25 páginasNuggets de PescadoCordova FernandoAinda não há avaliações
- Aguaymanto y Mango PTTTDocumento4 páginasAguaymanto y Mango PTTTCordova FernandoAinda não há avaliações
- 1351223018.transportadores de CadenasDocumento11 páginas1351223018.transportadores de CadenasEduardo Vidal CidAinda não há avaliações
- AISLADOSDocumento23 páginasAISLADOSCordova FernandoAinda não há avaliações
- Canaletas VibratoriasDocumento5 páginasCanaletas VibratoriasCordova FernandoAinda não há avaliações
- Ficha Tecnica de Productos Bilaminados y TrilaminadosDocumento1 páginaFicha Tecnica de Productos Bilaminados y TrilaminadosCordova FernandoAinda não há avaliações
- Aislado Proteico de QuinuaDocumento1 páginaAislado Proteico de QuinuaCordova FernandoAinda não há avaliações
- Variedades de UvaDocumento4 páginasVariedades de UvaCordova FernandoAinda não há avaliações
- BIORREACCIONDocumento1 páginaBIORREACCIONCordova FernandoAinda não há avaliações
- Vinos de ItaliaDocumento96 páginasVinos de ItaliaCordova Fernando100% (1)
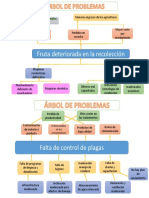
- Arbol de ProblemasDocumento2 páginasArbol de ProblemasCordova Fernando100% (2)
- BENEFICIOS y Aprovechamiento de La CarneDocumento3 páginasBENEFICIOS y Aprovechamiento de La CarneCordova FernandoAinda não há avaliações
- Industria de La Panificación WordDocumento24 páginasIndustria de La Panificación WordCordova FernandoAinda não há avaliações
- AISLADOSDocumento23 páginasAISLADOSCordova FernandoAinda não há avaliações
- Discusiones Conservas 1Documento2 páginasDiscusiones Conservas 1Cordova FernandoAinda não há avaliações
- Secuencia Didactica Tercero-La Diversidad de AnimalesDocumento19 páginasSecuencia Didactica Tercero-La Diversidad de AnimalesMagali Maldonado88% (17)
- AlacranDocumento8 páginasAlacranmiltonmartinezbAinda não há avaliações
- STARLINKDocumento2 páginasSTARLINKMario OrtaAinda não há avaliações
- Tarea II - Tarea III - Hormonas y PersonalidadDocumento16 páginasTarea II - Tarea III - Hormonas y PersonalidadHenrry Antigua GonzalezAinda não há avaliações
- Mapas Conceptuales - Seccion Xvii - XxiDocumento10 páginasMapas Conceptuales - Seccion Xvii - XxialezeyAinda não há avaliações
- Entendiendo Los Motores SumergiblesDocumento12 páginasEntendiendo Los Motores SumergiblesvicbtoAinda não há avaliações
- Analisis MatematicoDocumento12 páginasAnalisis MatematicoVeronica Martina Cacha Espinoza0% (1)
- Reporte de Alcances de Trabajo Celda ColumnaDocumento17 páginasReporte de Alcances de Trabajo Celda ColumnaJhon Stn Pecho IngaAinda não há avaliações
- Contenido Programático - Pensamiento Sistémico 2020-30 PDFDocumento8 páginasContenido Programático - Pensamiento Sistémico 2020-30 PDFLaura Gómez AmayaAinda não há avaliações
- SdraDocumento26 páginasSdracande rojasAinda não há avaliações
- Tesina Jorge Baños PDFDocumento264 páginasTesina Jorge Baños PDFpaulkohanAinda não há avaliações
- Buen Uso Del LaboratorioDocumento6 páginasBuen Uso Del Laboratoriobelen_reija_otero5038Ainda não há avaliações
- Ismael y La Salvacion de La TierraDocumento65 páginasIsmael y La Salvacion de La Tierrafabi_kuleff100% (1)
- Plan de Refuerzo ReligionDocumento2 páginasPlan de Refuerzo ReligionMonica Martinez50% (2)
- Emisiones de Las Formas 1Documento4 páginasEmisiones de Las Formas 1GabrielInsua100% (2)
- Los Alquimicos Jardines de BomarzoDocumento4 páginasLos Alquimicos Jardines de BomarzoRufino Fernandez Comendador100% (1)
- Canciones y Poesias A TacnaDocumento38 páginasCanciones y Poesias A TacnaYaneth Katherine Atamari Flores50% (6)
- Equilibrio IonicoDocumento23 páginasEquilibrio IonicoFroilan CanquiAinda não há avaliações
- 1 Minado, Transorte y Molienda Shery y XiomiDocumento10 páginas1 Minado, Transorte y Molienda Shery y XiomiGabriel Rodríguez SuttaAinda não há avaliações
- GlosarioDocumento3 páginasGlosarioGisel IvanniaAinda não há avaliações
- Traduccion Guias ESPEN de NE PDFDocumento3 páginasTraduccion Guias ESPEN de NE PDFDavid M. Lesmes100% (1)
- 85 Curiosidades Que Posiblemente No Conocias Sobre TintínDocumento6 páginas85 Curiosidades Que Posiblemente No Conocias Sobre TintínRamón Suárez50% (2)
- El Corto de Loja 24 08 2016Documento48 páginasEl Corto de Loja 24 08 2016JoséAndrésOrtizCuestaAinda não há avaliações
- Dinámica LinealDocumento3 páginasDinámica LinealjobishAinda não há avaliações
- Almendros Nestor - Dias de Una CamaraDocumento183 páginasAlmendros Nestor - Dias de Una CamaraDaniela MuñozAinda não há avaliações
- Mademsa Ventti 460 B Plus DryerDocumento14 páginasMademsa Ventti 460 B Plus DryerSusanaGulppiPintoAinda não há avaliações
- Unidad IVDocumento46 páginasUnidad IVAndrea Gerl8ngAinda não há avaliações
- Los Diagnósticos en La Infancia Alicia Stolkiner. PDF - CompressedDocumento27 páginasLos Diagnósticos en La Infancia Alicia Stolkiner. PDF - CompressedSara AdornoAinda não há avaliações
- Como Actua Dios en Los Eventos Casuales Juan Jose SanguinetiDocumento49 páginasComo Actua Dios en Los Eventos Casuales Juan Jose SanguinetiRicardo DiazAinda não há avaliações
- Autobús 7300-1Documento2 páginasAutobús 7300-1Alberto Caín100% (1)