Você também pode gostar
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5795)
- Review of Related Literature and StudiesDocumento10 páginasReview of Related Literature and StudiesJay-Rald Torres75% (4)
- Credit Card Data - Final Project Proposal - VictorDocumento1 páginaCredit Card Data - Final Project Proposal - VictorVictor KingbuilderAinda não há avaliações
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- BRM PaperDocumento9 páginasBRM PaperMalik Mughees AwanAinda não há avaliações
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Time Series Forecasting: Project Report On of Given Instrument BYDocumento29 páginasTime Series Forecasting: Project Report On of Given Instrument BYRahil RahmanAinda não há avaliações
- Contoh Interpretasi Data Pada SPSS 21Documento22 páginasContoh Interpretasi Data Pada SPSS 21Ahmad Fauzi NSAinda não há avaliações
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- The Use of Odonata Species For Environmental Assessment: A Meta-Analysis For The Neotropical RegionDocumento16 páginasThe Use of Odonata Species For Environmental Assessment: A Meta-Analysis For The Neotropical Regiongabriel vila verde nunesAinda não há avaliações
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Chapter 5 Discrete Probability DistributionsDocumento25 páginasChapter 5 Discrete Probability DistributionsGabrielaAinda não há avaliações
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- Factorial Design Notes and ExamplesDocumento20 páginasFactorial Design Notes and ExamplesAZ NdingwanAinda não há avaliações
- Assessment of The Suitability of Banana: (Musa Acuminata) Flour For Domestic FoodDocumento29 páginasAssessment of The Suitability of Banana: (Musa Acuminata) Flour For Domestic Foodedvin100% (2)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- Transforming Normal To Standard NormalDocumento14 páginasTransforming Normal To Standard NormalMarcy BoralAinda não há avaliações
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- StatistikinferialDocumento29 páginasStatistikinferialaditya meikoAinda não há avaliações
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Geostatistics For ApplicationDocumento151 páginasGeostatistics For ApplicationOlga100% (9)
- Dummy ChapterDocumento10 páginasDummy ChapterYared SolomonAinda não há avaliações
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- 53 IJFDE V2N3 PP 0053 Ink+dating 2w F PDFDocumento17 páginas53 IJFDE V2N3 PP 0053 Ink+dating 2w F PDFNikolaos KalantzisAinda não há avaliações
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- Best-Fit Probability Distributions and Return Periods, Northern CyprusDocumento10 páginasBest-Fit Probability Distributions and Return Periods, Northern CyprusNOURATEFNUMAN ALIJELAinda não há avaliações
- Lecture6 0Documento130 páginasLecture6 0hanoi6Ainda não há avaliações
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Chapter 11Documento30 páginasChapter 11Khang Lê VănAinda não há avaliações
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1091)
- 5 ErrorsDocumento21 páginas5 Errorsoliver senAinda não há avaliações
- Stationary and Nonstationary Series: T y y E T y S S T y T y S T y T y T yDocumento17 páginasStationary and Nonstationary Series: T y y E T y S S T y T y S T y T y T yProf B Sudhakar ReddyAinda não há avaliações
- Checked Batch 5a John Erroll O. Gesmundo 3bDocumento105 páginasChecked Batch 5a John Erroll O. Gesmundo 3bJan Jan HazeAinda não há avaliações
- Course Unit 6 - Z Test and T Test-2Documento11 páginasCourse Unit 6 - Z Test and T Test-2AnneDreaSamsonAinda não há avaliações
- MMGT6012 - Topic 7 - Time Series ForecastingDocumento46 páginasMMGT6012 - Topic 7 - Time Series ForecastingHuayu YangAinda não há avaliações
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- Fixed Effects, Random Effects Model Cheat SheetDocumento4 páginasFixed Effects, Random Effects Model Cheat SheetEric Rojas100% (1)
- Chapter 12Documento6 páginasChapter 12masrawy eduAinda não há avaliações
- F Distribution Table PDFDocumento2 páginasF Distribution Table PDFAdamAinda não há avaliações
- Sampling: Research Methodology: Tools, Methods and TechniquesDocumento39 páginasSampling: Research Methodology: Tools, Methods and TechniquesAmrezaa IskandarAinda não há avaliações
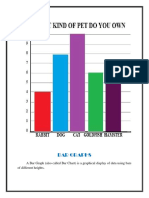
- Bar Graphs: A Bar Graph (Also Called Bar Chart) Is A Graphical Display of Data Using Bars of Different HeightsDocumento13 páginasBar Graphs: A Bar Graph (Also Called Bar Chart) Is A Graphical Display of Data Using Bars of Different HeightsMr 73ieAinda não há avaliações
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- PSPP TutorialDocumento24 páginasPSPP Tutorialyarrha100% (1)
- Senior High School TG LMDocumento5 páginasSenior High School TG LMAnalie CabanlitAinda não há avaliações
- Probability and Statistics (MMT-008) : Practical Assignment FileDocumento48 páginasProbability and Statistics (MMT-008) : Practical Assignment FileAnurag GargAinda não há avaliações