Você também pode gostar
- Metabolismo y efectos adversos de los CannabinoidesNo EverandMetabolismo y efectos adversos de los CannabinoidesAinda não há avaliações
- Caracteristicas de Las Fibras Lentas y RapidasDocumento4 páginasCaracteristicas de Las Fibras Lentas y RapidasFabian VascoAinda não há avaliações
- 13 Hemorragia Digestiva Alta No VaricosaDocumento9 páginas13 Hemorragia Digestiva Alta No VaricosaJoel GuishaAinda não há avaliações
- Biomarcadores EpigenéticosDocumento28 páginasBiomarcadores EpigenéticosCarlos DiezAinda não há avaliações
- Causas de Lesion CelularDocumento10 páginasCausas de Lesion CelularAna HernándezAinda não há avaliações
- Cardiopatias Congenitas CianoticasDocumento35 páginasCardiopatias Congenitas CianoticasAngélica Huerta100% (1)
- EspirometríaDocumento17 páginasEspirometríaMarianela Vásquez Araya100% (1)
- Estructura Del ADNDocumento22 páginasEstructura Del ADNJonatan DuranAinda não há avaliações
- La Condición Previa Del PacienteDocumento4 páginasLa Condición Previa Del PacienteAlyssa GarciaAinda não há avaliações
- Reflejo Extensor Cruzado 1Documento5 páginasReflejo Extensor Cruzado 1DANIEL ALEJANDROAinda não há avaliações
- Trifolio EspaciadorDocumento2 páginasTrifolio EspaciadorAndré Ponce BautistaAinda não há avaliações
- Gu - A Uso InhaladoresDocumento35 páginasGu - A Uso InhaladoresCarlos CalvacheAinda não há avaliações
- Reflejo MiotáticoDocumento1 páginaReflejo MiotáticoEdwin AlexanderAinda não há avaliações
- Cuadro Comparativo CardiologiaDocumento8 páginasCuadro Comparativo CardiologiaSusy SilvaAinda não há avaliações
- Corticoides TopicosDocumento2 páginasCorticoides TopicosBe AMAinda não há avaliações
- CotransmisionDocumento19 páginasCotransmisionGabyta Paladines YangeAinda não há avaliações
- Cristales de Charcot Leyden Definicion 8814 Mxe6stDocumento1 páginaCristales de Charcot Leyden Definicion 8814 Mxe6stZoraide OrtegamAinda não há avaliações
- Antibioticos CuadrosDocumento10 páginasAntibioticos CuadrosJulio SilvaAinda não há avaliações
- Divertículo de MeckelDocumento5 páginasDivertículo de MeckelDaynor E. Guzman VeraAinda não há avaliações
- Sindrome de Morris 2020Documento6 páginasSindrome de Morris 2020leslyAinda não há avaliações
- Sindrome de MorrisDocumento2 páginasSindrome de MorrisJorge OrtizAinda não há avaliações
- Neuro TrasmisoresDocumento5 páginasNeuro Trasmisorescatalina acosta100% (1)
- Edema Agudo de PulmonDocumento11 páginasEdema Agudo de PulmonkeyberliAinda não há avaliações
- Icc Semiologia.Documento30 páginasIcc Semiologia.Cristóbal Graterol BarriosAinda não há avaliações
- HipolipemiantesDocumento21 páginasHipolipemiantesKelly SVAinda não há avaliações
- Activación LinfocitariaDocumento5 páginasActivación LinfocitariaDavidZuñigaPorfirioAinda não há avaliações
- SINDROME de MORRISDocumento1 páginaSINDROME de MORRISAnaAinda não há avaliações
- Síndrome de Insensibilidad de Los AndrógenosDocumento17 páginasSíndrome de Insensibilidad de Los AndrógenoshevelinAinda não há avaliações
- Ejercicios de Gasometría - 2Documento2 páginasEjercicios de Gasometría - 2Denise GuerreroAinda não há avaliações
- Fisipopatologia HipercapniaDocumento5 páginasFisipopatologia HipercapniaPaulina Rubilar FerisAinda não há avaliações
- Fármacos AntirreumáticosDocumento24 páginasFármacos Antirreumáticosluis1805100% (1)
- Uso de AerocamaraDocumento1 páginaUso de AerocamaraMaria Paz Díaz HerreraAinda não há avaliações
- BorreliaDocumento12 páginasBorreliaGabriela VHAinda não há avaliações
- HIPERCAPNIADocumento5 páginasHIPERCAPNIARuttiiErazooAinda não há avaliações
- A Reflejo MiotaticoDocumento13 páginasA Reflejo MiotaticoAlberto TorresAinda não há avaliações
- Informe AnticoagulantesDocumento27 páginasInforme AnticoagulantesJavieraPssAinda não há avaliações
- Enfermedades de La CoagulaciónDocumento12 páginasEnfermedades de La CoagulaciónAlan Andree Rengifo ArevaloAinda não há avaliações
- Tromboembolismo Pulmonar: "T" HelperDocumento10 páginasTromboembolismo Pulmonar: "T" HelperJuan Carlos AguilarAinda não há avaliações
- Estructura BacterianaDocumento29 páginasEstructura BacterianaSandra KattAinda não há avaliações
- HISTORIA NATURAL de Neumonia Esquema Copia 1Documento2 páginasHISTORIA NATURAL de Neumonia Esquema Copia 1Susana GuerraAinda não há avaliações
- EXPO PenicilinasDocumento43 páginasEXPO PenicilinasBilo Orw100% (1)
- Arterioesclerosis PM1Documento37 páginasArterioesclerosis PM1GOPMY100% (1)
- Trastornos de La SexualidadDocumento17 páginasTrastornos de La SexualidadGabo BurrolaAinda não há avaliações
- Presiones en El Sistema PulmonarDocumento11 páginasPresiones en El Sistema PulmonarCitlali GarciaAinda não há avaliações
- Patogenicidad PDFDocumento61 páginasPatogenicidad PDFLaboratoriosClinicosZaragozaAinda não há avaliações
- Resumen InmunologíaDocumento25 páginasResumen InmunologíaAli Partiido100% (1)
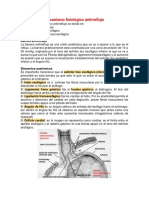
- Mecanismo Fisiológico AntirreflujoDocumento14 páginasMecanismo Fisiológico AntirreflujoKendra TelloAinda não há avaliações
- Apuntes 3er Parcial URODocumento99 páginasApuntes 3er Parcial UROAlejandro ÁlvarezAinda não há avaliações
- Histamina y AntihistamínicosDocumento36 páginasHistamina y AntihistamínicosLuis Daniel Caudillo RodriguezAinda não há avaliações
- Gooman Cap 48 Principios Generales Del Tratamiento AntimicrobianoDocumento7 páginasGooman Cap 48 Principios Generales Del Tratamiento AntimicrobianoMiguel PalominoAinda não há avaliações
- Fo-Ter-04 Folleto Inhalocámaras - InhaladorDocumento2 páginasFo-Ter-04 Folleto Inhalocámaras - InhaladorEmerson JaimesAinda não há avaliações
- Seminario Hepatitis BDocumento16 páginasSeminario Hepatitis BDavid Sanchez Juarez100% (1)
- AMEBIASISDocumento7 páginasAMEBIASISRomi Scarlet CanedoAinda não há avaliações
- TabaquismoDocumento15 páginasTabaquismoMonicaFerradisAinda não há avaliações
- Penicilin AsDocumento32 páginasPenicilin As4317322100% (1)
- Neumonia Asociada A VentiladorDocumento15 páginasNeumonia Asociada A VentiladorLuis Lopez100% (1)
- ANGIOLOGIADocumento18 páginasANGIOLOGIAverona3010Ainda não há avaliações
- Laminario de PIEL Y ANEXOS. LENYS SOLORZANO.Documento5 páginasLaminario de PIEL Y ANEXOS. LENYS SOLORZANO.lenys solorzanoAinda não há avaliações
- Antivirales 1Documento25 páginasAntivirales 1Polaco54100% (1)
- Bases CarcinogenesisDocumento75 páginasBases Carcinogenesiscit.valdez100% (9)
- Generalidades CancerDocumento9 páginasGeneralidades CancerChipiRosalesAinda não há avaliações
- Bibliografias BlogDocumento1 páginaBibliografias BlogChipiRosalesAinda não há avaliações
- Reti No Blast OmaDocumento3 páginasReti No Blast OmaChipiRosalesAinda não há avaliações
- Causas Generales de CancerDocumento6 páginasCausas Generales de CancerChipiRosalesAinda não há avaliações
- Generalidades CancerDocumento9 páginasGeneralidades CancerChipiRosalesAinda não há avaliações
- Oxansil 17 PDFDocumento1 páginaOxansil 17 PDFAspro Brazil Se IglesiasAinda não há avaliações
- TESIS RECOPILACION SORIANO DDocumento4 páginasTESIS RECOPILACION SORIANO DPiero SorianoAinda não há avaliações
- Informe de Identificacion de Carbon, Hidrogeno, Nitrogeno y HalogenosDocumento3 páginasInforme de Identificacion de Carbon, Hidrogeno, Nitrogeno y HalogenosAlejandro RendonAinda não há avaliações
- Extincion de La Respuesta CondicionadaDocumento12 páginasExtincion de La Respuesta Condicionadayovana aguilar ceronAinda não há avaliações
- Laboratorio de Edafologia 9 SalinidadDocumento4 páginasLaboratorio de Edafologia 9 SalinidadJeferDavila0% (1)
- MarcoometDocumento8 páginasMarcoometXIOMARA DEL CARMEN BARRANZUELA NEYRAAinda não há avaliações
- August 2017 Tool News - SpanishDocumento8 páginasAugust 2017 Tool News - SpanishCarlos GajardoAinda não há avaliações
- Diario de SueñosDocumento2 páginasDiario de SueñosAnonymous e3vAkC9IZ5Ainda não há avaliações
- Riesgo PublicoDocumento16 páginasRiesgo PublicoSamir Vargas ReyesAinda não há avaliações
- Evaluación Del AguaDocumento4 páginasEvaluación Del AguaBarbara JaraAinda não há avaliações
- Estimulación TempranaDocumento57 páginasEstimulación TempranaAndres JaramilloAinda não há avaliações
- CÓMO SE CRIABA A LOS NIÑOS EN LA ANTIGUA GRECIA - Docx TÉCNICASDocumento2 páginasCÓMO SE CRIABA A LOS NIÑOS EN LA ANTIGUA GRECIA - Docx TÉCNICASJhon JimenezAinda não há avaliações
- Riesgos en La Industria TextilDocumento7 páginasRiesgos en La Industria TextilAmparo MoraAinda não há avaliações
- VPH - Pablo Campos PDFDocumento22 páginasVPH - Pablo Campos PDFLaura Perez ZarazúaAinda não há avaliações
- FarmacologiaDocumento32 páginasFarmacologiaOscar VejarAinda não há avaliações
- Ensayo Sobre La Aplicacion de Quimica en La IndustriaDocumento11 páginasEnsayo Sobre La Aplicacion de Quimica en La Industriajhudith torresAinda não há avaliações
- Libro - Edificaciones de Mamposteria para ViviendaDocumento607 páginasLibro - Edificaciones de Mamposteria para Viviendajuan romero agustin67% (3)
- Divisiones de La Anatomía y FisiologíaDocumento10 páginasDivisiones de La Anatomía y FisiologíaangelesAinda não há avaliações
- Exp. 4378-2018-92Documento12 páginasExp. 4378-2018-92haiden de la palmaAinda não há avaliações
- Proyecto Escabeche - Esmeralda CortesDocumento16 páginasProyecto Escabeche - Esmeralda CortesAlcatraz CortesAinda não há avaliações
- Cadena Alimentaria PDFDocumento1 páginaCadena Alimentaria PDFMaría Salazar CastroAinda não há avaliações
- Examen TS LDocumento36 páginasExamen TS LmaryteAinda não há avaliações
- Contrato de ResiduosDocumento6 páginasContrato de ResiduosRodrigo Vidal EscrivàAinda não há avaliações
- 1 TDR Camara de Empalme y MedicionDocumento23 páginas1 TDR Camara de Empalme y MedicionAnthony Soto MegoAinda não há avaliações
- Envases de PapelDocumento38 páginasEnvases de Papelsmanuelo362Ainda não há avaliações
- Fertilizacion Calcica ManiDocumento12 páginasFertilizacion Calcica ManiAndres VbAinda não há avaliações
- Guias de AdultoDocumento4 páginasGuias de AdultoYael Cedeño OlivoAinda não há avaliações
- La Anomalía o Alteración Psíquica en La Interpretación JurisprudencialDocumento33 páginasLa Anomalía o Alteración Psíquica en La Interpretación JurisprudencialKikoCalvin100% (2)
- Locura 2Documento16 páginasLocura 2Kevin PozoAinda não há avaliações
- Soldadura de Geomenbrana Por ExtrusionDocumento3 páginasSoldadura de Geomenbrana Por ExtrusionKevin Shinder Jesus Colque EscobedoAinda não há avaliações