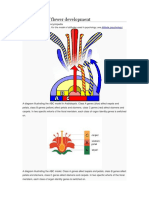
Você também pode gostar
- Alexander StainDocumento8 páginasAlexander StainpantremenoAinda não há avaliações
- tmp684 TMPDocumento6 páginastmp684 TMPFrontiersAinda não há avaliações
- The Effects of Cross & Self-Fertilisation in the Vegetable KingdomNo EverandThe Effects of Cross & Self-Fertilisation in the Vegetable KingdomAinda não há avaliações
- BARRET 98 Evolution of Mating StrategiesDocumento7 páginasBARRET 98 Evolution of Mating StrategiesCommon CannaAinda não há avaliações
- Flowers Chap17Documento15 páginasFlowers Chap17Cheret LengAinda não há avaliações
- Das 2016Documento38 páginasDas 2016rismaAinda não há avaliações
- M V. C S A. K: Orrichia Frutescens and - ArborescensDocumento10 páginasM V. C S A. K: Orrichia Frutescens and - Arborescensnica_salvadorAinda não há avaliações
- ABC Model of Flower DevelopmentDocumento11 páginasABC Model of Flower DevelopmentAlexandra Maria NeaguAinda não há avaliações
- Stamen Structure and Function: R. J. Scott, M. Spielman, and H. G. DickinsonDocumento15 páginasStamen Structure and Function: R. J. Scott, M. Spielman, and H. G. DickinsonAJMRAinda não há avaliações
- Palm 2016Documento13 páginasPalm 2016Boran KzAinda não há avaliações
- Project Shia College FinalDocumento42 páginasProject Shia College FinalNeeraj KaushalAinda não há avaliações
- Insperata (Poaceae) : Comparative Results For Allozymes andDocumento9 páginasInsperata (Poaceae) : Comparative Results For Allozymes andSultan Ahmad KhanAinda não há avaliações
- Dun Well 2010Documento48 páginasDun Well 2010lacisagAinda não há avaliações
- British Ecological Society Journal of Ecology: This Content Downloaded From 200.24.16.65 On Sat, 08 Jun 2019 16:36:43 UTCDocumento13 páginasBritish Ecological Society Journal of Ecology: This Content Downloaded From 200.24.16.65 On Sat, 08 Jun 2019 16:36:43 UTCJESSICA HERRERA SANCHEZAinda não há avaliações
- Muchhala 2007Documento11 páginasMuchhala 2007DANIEL POSADA GUTIERREZAinda não há avaliações
- Non-Pollinator Agents of SelectionDocumento21 páginasNon-Pollinator Agents of SelectionAntonio mlnAinda não há avaliações
- BiochemistryDocumento14 páginasBiochemistryasish MohapatraAinda não há avaliações
- Molecular Mechanisms of Flower Development: An Armchair GuideDocumento11 páginasMolecular Mechanisms of Flower Development: An Armchair GuideCarina JLAinda não há avaliações
- The Role of Male Flowers in Andromonoecious Species: Energetic Costs and Siring Success in SolanumDocumento9 páginasThe Role of Male Flowers in Andromonoecious Species: Energetic Costs and Siring Success in SolanumArya GuganAinda não há avaliações
- Principles of Genetics: Module - 3Documento26 páginasPrinciples of Genetics: Module - 3Indrojyoti MondalAinda não há avaliações
- Development: Flowering TransitionDocumento3 páginasDevelopment: Flowering TransitionNovember RolfAinda não há avaliações
- Rayburn Genome SizeDocumento6 páginasRayburn Genome SizeSebastian DavisAinda não há avaliações
- Martén - Evolutionary Breakdown of Pollination SpecializationDocumento15 páginasMartén - Evolutionary Breakdown of Pollination SpecializationRodrigo CarralAinda não há avaliações
- Robles 2005 ABC ModelDocumento11 páginasRobles 2005 ABC ModelJegan Passiflora EdulisAinda não há avaliações
- Self Incompatibility 2012 InroadsDocumento12 páginasSelf Incompatibility 2012 InroadsGulzar SangheraAinda não há avaliações
- The Use of Molecular Markers To Analyze The Inheritance of Morphological and Developmental Traits in AppleDocumento6 páginasThe Use of Molecular Markers To Analyze The Inheritance of Morphological and Developmental Traits in AppleFernando Castro EchavezAinda não há avaliações
- Pomegranate Flower TypeDocumento10 páginasPomegranate Flower TypeAki FujikawaAinda não há avaliações
- A Global Test of The Pollination Syndrome HypothesisDocumento10 páginasA Global Test of The Pollination Syndrome HypothesisConstanza Andía AvendañoAinda não há avaliações
- Bisexual & Male Flower PDFDocumento10 páginasBisexual & Male Flower PDFIcsAinda não há avaliações
- Brun Et Al. - 2018 - Seed Germination in Parasitic Plants What InsightDocumento16 páginasBrun Et Al. - 2018 - Seed Germination in Parasitic Plants What InsightJustin PawlonskiAinda não há avaliações
- PitahayaDocumento11 páginasPitahayajorgeAinda não há avaliações
- BIOS606 2012 Module 01Documento13 páginasBIOS606 2012 Module 01hlch20011973Ainda não há avaliações
- Botanical Society of AmericaDocumento11 páginasBotanical Society of AmericaAaron MeloAinda não há avaliações
- Heterosis: A 1 A A B CDocumento9 páginasHeterosis: A 1 A A B CpraveenAinda não há avaliações
- MBU Genetic Control of FlowersDocumento4 páginasMBU Genetic Control of FlowersketakeeAinda não há avaliações
- Karakteristik Bunga TelangDocumento7 páginasKarakteristik Bunga TelangRahmahPuspitaSariAinda não há avaliações
- Phylogeny and Diversification of BryophytesDocumento26 páginasPhylogeny and Diversification of BryophytesHamengku AgengAinda não há avaliações
- Shaw, J. & K. Renzaglia. (2004) - Phylogeny and Diversification of Bryophytes. American PDFDocumento26 páginasShaw, J. & K. Renzaglia. (2004) - Phylogeny and Diversification of Bryophytes. American PDFDiego Andres Moreno GaonaAinda não há avaliações
- Sun Et Al. 2011 - Why Do Stigmas Move in A Flexistylous Plant (QM)Documento8 páginasSun Et Al. 2011 - Why Do Stigmas Move in A Flexistylous Plant (QM)Marcelo MorettoAinda não há avaliações
- MSc. Lecture 1Documento14 páginasMSc. Lecture 1michael agutoAinda não há avaliações
- ABC Model of Flower Development - WikipediaDocumento9 páginasABC Model of Flower Development - WikipediaCarina JLAinda não há avaliações
- Scientific Article SummaryDocumento5 páginasScientific Article Summaryapi-325188603Ainda não há avaliações
- QuesadaETAL2008 FullDocumento12 páginasQuesadaETAL2008 FullSilviyaniAinda não há avaliações
- ATLAN Et Al-2010-Journal of Evolutionary BiologyDocumento10 páginasATLAN Et Al-2010-Journal of Evolutionary BiologyDarielaNegrínPérezAinda não há avaliações
- PDF PDFDocumento9 páginasPDF PDFMartín Rojas TorrejónAinda não há avaliações
- Pathogenic FungiDocumento2 páginasPathogenic FungiLeda Rodriguez PossoAinda não há avaliações
- Flower DevelopmentDocumento12 páginasFlower DevelopmentthenderingAinda não há avaliações
- Bunga 4Documento2 páginasBunga 4yosephineikaaAinda não há avaliações
- Phylogenetic Distribution and Evolution of MycorrhizasDocumento65 páginasPhylogenetic Distribution and Evolution of MycorrhizasUtanka DeAinda não há avaliações
- Sex Chromosome in PlantsDocumento32 páginasSex Chromosome in PlantsAshutosh MukherjeeAinda não há avaliações
- Spore Fitness Components Do Not Differ Between Diploid and Allotetraploid Species of Dryopteris (Dryopteridaceae)Documento10 páginasSpore Fitness Components Do Not Differ Between Diploid and Allotetraploid Species of Dryopteris (Dryopteridaceae)Neni InayahAinda não há avaliações
- Plcell v1 1 37Documento16 páginasPlcell v1 1 37HaienasrAinda não há avaliações
- Kwak Et Al. 2012Documento8 páginasKwak Et Al. 2012Gisele HolandaAinda não há avaliações
- 1 SM PDFDocumento12 páginas1 SM PDFCarlos Lesmes DíazAinda não há avaliações
- Bogarin - 2016 - Orquidea EpidendrumDocumento34 páginasBogarin - 2016 - Orquidea EpidendrumDavid AngaritaAinda não há avaliações
- Functional Gynodioecy in Opuntia Quimilo (Cactaceae), A Tree Cactus Pollinated by Bees and HummingbirdsDocumento9 páginasFunctional Gynodioecy in Opuntia Quimilo (Cactaceae), A Tree Cactus Pollinated by Bees and HummingbirdsOrlando Luis GarcíaAinda não há avaliações
- Sdarticle 019Documento3 páginasSdarticle 019Elsa Fernita ManullangAinda não há avaliações
- Daniel&SchoenDocumento10 páginasDaniel&SchoenHerniqueAinda não há avaliações
- Genetic and Morphological Differentiation Patterns Between Sister Species: The Case of Onthophagus Taurus and Onthophagus IllyricusDocumento15 páginasGenetic and Morphological Differentiation Patterns Between Sister Species: The Case of Onthophagus Taurus and Onthophagus IllyricusJulio SubagioAinda não há avaliações
- DeQueiroz.2007 Species Concepts and Species DelimitationDocumento8 páginasDeQueiroz.2007 Species Concepts and Species DelimitationJulio SubagioAinda não há avaliações
- Bios116-Drosophila Eye PigmentDocumento3 páginasBios116-Drosophila Eye PigmentJulio SubagioAinda não há avaliações
- DeQueiroz.2005.Different Species Problems and Their ResolutionDocumento7 páginasDeQueiroz.2005.Different Species Problems and Their ResolutionJulio SubagioAinda não há avaliações
- Your Thought and MineDocumento2 páginasYour Thought and MineJulio SubagioAinda não há avaliações
- Ea MsdsDocumento6 páginasEa Msdsapi-263411629Ainda não há avaliações
- Bios116-Drosophila Eye PigmentDocumento3 páginasBios116-Drosophila Eye PigmentJulio SubagioAinda não há avaliações
- Ea MsdsDocumento6 páginasEa Msdsapi-263411629Ainda não há avaliações
- Ea MsdsDocumento6 páginasEa Msdsapi-263411629Ainda não há avaliações
- 04 - System, Matter & EnergyDocumento14 páginas04 - System, Matter & EnergyJulio SubagioAinda não há avaliações
- Ea MsdsDocumento6 páginasEa Msdsapi-263411629Ainda não há avaliações
- 03 Critical ThinkingDocumento11 páginas03 Critical ThinkingJulio SubagioAinda não há avaliações
- 04 - System, Matter & EnergyDocumento14 páginas04 - System, Matter & EnergyJulio SubagioAinda não há avaliações
- 03 Critical ThinkingDocumento11 páginas03 Critical ThinkingJulio SubagioAinda não há avaliações
- 188 Integrated Pest Management: Current and Future StrategiesDocumento10 páginas188 Integrated Pest Management: Current and Future StrategiesTaufik RizkiandiAinda não há avaliações
- Why Not Leeks?Documento2 páginasWhy Not Leeks?Julio SubagioAinda não há avaliações
- Why Not Leeks?Documento2 páginasWhy Not Leeks?Julio SubagioAinda não há avaliações
- Why Not Leeks?Documento2 páginasWhy Not Leeks?Julio SubagioAinda não há avaliações
- The Medicinal Plants of TheDocumento180 páginasThe Medicinal Plants of TheRein Alcira100% (2)
- Gesc 112Documento10 páginasGesc 112Kalpavriksha1974Ainda não há avaliações
- CH 2 AssignmentDocumento7 páginasCH 2 AssignmentRitu BarthwalAinda não há avaliações
- Crsci 1100 Exercise No.5Documento4 páginasCrsci 1100 Exercise No.5Angelyn CayabanAinda não há avaliações
- Hybrid Seed Production of CucurbitsDocumento6 páginasHybrid Seed Production of CucurbitsMd. Sajjad HossenAinda não há avaliações
- Vol. XIDocumento333 páginasVol. XICamila AlcantaraAinda não há avaliações
- Ipil IpilDocumento4 páginasIpil IpilJohn Rudolf CatalanAinda não há avaliações
- Plant Classification DegreeDocumento57 páginasPlant Classification Degreejohnsonsamuel090677746Ainda não há avaliações
- Aipmt 17 18 XII Bot Study Package 4 SET 1 Chapter 1Documento16 páginasAipmt 17 18 XII Bot Study Package 4 SET 1 Chapter 1Riya Mishra100% (3)
- Mohan Ram STS 1982 by HTTPDocumento7 páginasMohan Ram STS 1982 by HTTPmurciano207100% (2)
- I. A. Table 1: Flower Parts Illustration Example: Systematics Act. 3 Phytography: FlowerDocumento19 páginasI. A. Table 1: Flower Parts Illustration Example: Systematics Act. 3 Phytography: FlowerAngeline Casis CordovaAinda não há avaliações
- Subject-Biology Chapter - 5 Morphology of Flowering PlantsDocumento35 páginasSubject-Biology Chapter - 5 Morphology of Flowering PlantsNikunj kumar chaurasiyaAinda não há avaliações
- Mode of ReproductionDocumento33 páginasMode of ReproductionMd. Rayhan Ali RejviAinda não há avaliações
- CLASS - 7th SCIENCE, CHAPTER 8 (REPRODUCTION IN PLANTSDocumento1 páginaCLASS - 7th SCIENCE, CHAPTER 8 (REPRODUCTION IN PLANTSexperimentoboyAinda não há avaliações
- CBSE Class 10 Science Revision Notes Chapter - 8 How Do Organism ReproduceDocumento10 páginasCBSE Class 10 Science Revision Notes Chapter - 8 How Do Organism ReproduceAVIRUP DASAinda não há avaliações
- Flowers 121024072042 Phpapp02Documento42 páginasFlowers 121024072042 Phpapp02doncer100% (1)
- Velarde Exercise4 PhytographyDocumento104 páginasVelarde Exercise4 PhytographytolisAinda não há avaliações
- Acp 301 Arable Crop Production 1Documento46 páginasAcp 301 Arable Crop Production 1JayjayAinda não há avaliações
- Breeding Field Crop by Pohlman 5th EdDocumento501 páginasBreeding Field Crop by Pohlman 5th EdQurat ul AinAinda não há avaliações
- 2017 Pollinationin Roses Elsevier Modulein Life Sciences 050703Documento6 páginas2017 Pollinationin Roses Elsevier Modulein Life Sciences 050703Cricket with AliAinda não há avaliações
- Morphology PremedDocumento50 páginasMorphology PremedAASHISHAinda não há avaliações
- Bangladesh Seed Production PDFDocumento125 páginasBangladesh Seed Production PDFYanoAinda não há avaliações
- Class: 10 BiologyDocumento12 páginasClass: 10 BiologyRehan PervaizAinda não há avaliações
- How Do Organism ReproduceDocumento40 páginasHow Do Organism ReproduceNakshatra PaliwalAinda não há avaliações
- Chapter 2: Biology of Agricultural CropsDocumento8 páginasChapter 2: Biology of Agricultural CropsDwight Luciano T MendezAinda não há avaliações
- Flowering PlantsDocumento17 páginasFlowering Plantsjoshua.brussell100% (2)
- 5-Morphology of Flowering PlantsDocumento15 páginas5-Morphology of Flowering PlantsVasant Kumar VarmaAinda não há avaliações
- Angiosperm PDFDocumento2 páginasAngiosperm PDFLiza Mortel CoralesAinda não há avaliações
- Reproductive Systems in PlantsDocumento32 páginasReproductive Systems in PlantsDaud ObalolaAinda não há avaliações
- Xam Idea Biology Class 12Documento517 páginasXam Idea Biology Class 12ssshuuushhhAinda não há avaliações
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNo EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisNota: 3.5 de 5 estrelas3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNo EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceNota: 4.5 de 5 estrelas4.5/5 (516)
- Tales from Both Sides of the Brain: A Life in NeuroscienceNo EverandTales from Both Sides of the Brain: A Life in NeuroscienceNota: 3 de 5 estrelas3/5 (18)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)No EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Nota: 4 de 5 estrelas4/5 (378)
- Gut: the new and revised Sunday Times bestsellerNo EverandGut: the new and revised Sunday Times bestsellerNota: 4 de 5 estrelas4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityNo EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityNota: 4 de 5 estrelas4/5 (3)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNo EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsNota: 4.5 de 5 estrelas4.5/5 (6)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouNo EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouNota: 4.5 de 5 estrelas4.5/5 (62)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeNo EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeAinda não há avaliações
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNo EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperNota: 4.5 de 5 estrelas4.5/5 (15)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNo EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesNota: 4.5 de 5 estrelas4.5/5 (397)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNo EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionNota: 4 de 5 estrelas4/5 (811)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldNo EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldNota: 4 de 5 estrelas4/5 (595)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNo Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessNota: 4 de 5 estrelas4/5 (33)
- Who's in Charge?: Free Will and the Science of the BrainNo EverandWho's in Charge?: Free Will and the Science of the BrainNota: 4 de 5 estrelas4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorAinda não há avaliações
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindNo EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindNota: 4.5 de 5 estrelas4.5/5 (93)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedNo EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedNota: 4 de 5 estrelas4/5 (11)
- Good Without God: What a Billion Nonreligious People Do BelieveNo EverandGood Without God: What a Billion Nonreligious People Do BelieveNota: 4 de 5 estrelas4/5 (66)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemNo EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemNota: 4.5 de 5 estrelas4.5/5 (115)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldNo EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldNota: 4.5 de 5 estrelas4.5/5 (18)
- Seven and a Half Lessons About the BrainNo EverandSeven and a Half Lessons About the BrainNota: 4 de 5 estrelas4/5 (109)
- Human: The Science Behind What Makes Your Brain UniqueNo EverandHuman: The Science Behind What Makes Your Brain UniqueNota: 3.5 de 5 estrelas3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildNo EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildNota: 4.5 de 5 estrelas4.5/5 (44)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessNo EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessNota: 4 de 5 estrelas4/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondNo EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondNota: 4 de 5 estrelas4/5 (4)