Você também pode gostar
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Green and White Zero Waste Living Education Video PresentationDocumento12 páginasGreen and White Zero Waste Living Education Video PresentationNicole SarileAinda não há avaliações
- Human Development IndexDocumento17 páginasHuman Development IndexriyaAinda não há avaliações
- Frbiblio PDFDocumento16 páginasFrbiblio PDFreolox100% (1)
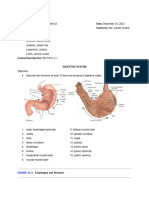
- Digestive System LabsheetDocumento4 páginasDigestive System LabsheetKATHLEEN MAE HERMOAinda não há avaliações
- AITAS 8th Doctor SourcebookDocumento192 páginasAITAS 8th Doctor SourcebookClaudio Caceres100% (13)
- Hankinson - Location Branding - A Study of The Branding Practices of 12 English CitiesDocumento16 páginasHankinson - Location Branding - A Study of The Branding Practices of 12 English CitiesNatalia Ney100% (1)
- ICTSAS601 Student Assessment Tasks 2020Documento30 páginasICTSAS601 Student Assessment Tasks 2020Lok SewaAinda não há avaliações
- Tesmec Catalogue TmeDocumento208 páginasTesmec Catalogue TmeDidier solanoAinda não há avaliações
- 4as Lesson PlanDocumento10 páginas4as Lesson PlanMannuelle Gacud100% (2)
- User Manual For Inquisit's Attentional Network TaskDocumento5 páginasUser Manual For Inquisit's Attentional Network TaskPiyush ParimooAinda não há avaliações
- Quiz 07Documento15 páginasQuiz 07Ije Love100% (1)
- PS4 ListDocumento67 páginasPS4 ListAnonymous yNw1VyHAinda não há avaliações
- Fort - Fts - The Teacher and ¿Mommy Zarry AdaptaciónDocumento90 páginasFort - Fts - The Teacher and ¿Mommy Zarry AdaptaciónEvelin PalenciaAinda não há avaliações
- NI 43-101 Technical Report - Lithium Mineral Resource Estimate Zeus Project, Clayton Valley, USADocumento71 páginasNI 43-101 Technical Report - Lithium Mineral Resource Estimate Zeus Project, Clayton Valley, USAGuillaume De SouzaAinda não há avaliações
- (Adolescence and Education) Tim Urdan, Frank Pajares - Academic Motivation of Adolescents-IAP - Information Age Publishing (2004) PDFDocumento384 páginas(Adolescence and Education) Tim Urdan, Frank Pajares - Academic Motivation of Adolescents-IAP - Information Age Publishing (2004) PDFAllenAinda não há avaliações
- Parts Catalog: TJ053E-AS50Documento14 páginasParts Catalog: TJ053E-AS50Andre FilipeAinda não há avaliações
- 5g-core-guide-building-a-new-world Переход от лте к 5г английскийDocumento13 páginas5g-core-guide-building-a-new-world Переход от лте к 5г английскийmashaAinda não há avaliações
- Filehost - CIA - Mind Control Techniques - (Ebook 197602 .TXT) (TEC@NZ)Documento52 páginasFilehost - CIA - Mind Control Techniques - (Ebook 197602 .TXT) (TEC@NZ)razvan_9100% (1)
- Lecture 4 PDFDocumento9 páginasLecture 4 PDFVarun SinghalAinda não há avaliações
- PUERPERAL SEPSIS CoverDocumento9 páginasPUERPERAL SEPSIS CoverKerpersky LogAinda não há avaliações
- Review Questions & Answers For Midterm1: BA 203 - Financial Accounting Fall 2019-2020Documento11 páginasReview Questions & Answers For Midterm1: BA 203 - Financial Accounting Fall 2019-2020Ulaş GüllenoğluAinda não há avaliações
- Dragon Ball AbrigedDocumento8 páginasDragon Ball AbrigedAlexander SusmanAinda não há avaliações
- Foundations For Assisting in Home Care 1520419723Documento349 páginasFoundations For Assisting in Home Care 1520419723amasrurAinda não há avaliações
- CS 124/LINGUIST 180 From Languages To Information: Conversational AgentsDocumento58 páginasCS 124/LINGUIST 180 From Languages To Information: Conversational AgentsamanAinda não há avaliações
- SRL CompressorsDocumento20 páginasSRL Compressorssthe03Ainda não há avaliações
- Introduction To Astronomical PhotometryDocumento452 páginasIntroduction To Astronomical PhotometrySergio Alejandro Fuentealba ZuñigaAinda não há avaliações
- Allen F. y D. Gale. Comparative Financial SystemsDocumento80 páginasAllen F. y D. Gale. Comparative Financial SystemsCliffordTorresAinda não há avaliações
- Simplified Electronic Design of The Function : ARMTH Start & Stop SystemDocumento6 páginasSimplified Electronic Design of The Function : ARMTH Start & Stop SystembadrAinda não há avaliações
- Placement TestDocumento6 páginasPlacement TestNovia YunitazamiAinda não há avaliações
- Picc Lite ManualDocumento366 páginasPicc Lite Manualtanny_03Ainda não há avaliações