Você também pode gostar
- C134 1945-1960 PDFDocumento16 páginasC134 1945-1960 PDFstein godoy pachecoAinda não há avaliações
- Control of Sleep and WakefulnessDocumento101 páginasControl of Sleep and WakefulnessLevente BalázsAinda não há avaliações
- Rushil Surapaneni January 18, 2017 Psych 133 Lecture Notes: ST NDDocumento7 páginasRushil Surapaneni January 18, 2017 Psych 133 Lecture Notes: ST NDRushil SurapaneniAinda não há avaliações
- Physiology of Normal Sleep: From Young To Old: V. Mohan KumarDocumento11 páginasPhysiology of Normal Sleep: From Young To Old: V. Mohan KumarRosalinaAinda não há avaliações
- The Sleep SwitchDocumento6 páginasThe Sleep SwitchtomasdjAinda não há avaliações
- The Neurobiology of DreamingDocumento12 páginasThe Neurobiology of DreamingmehrasAinda não há avaliações
- Neurobiology of Sleep: Madhu Kalia4Documento5 páginasNeurobiology of Sleep: Madhu Kalia4Julian ReyesAinda não há avaliações
- Physiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationDocumento10 páginasPhysiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationMukernas Perdossi 2021Ainda não há avaliações
- Stenberg 2007Documento18 páginasStenberg 2007NikeAinda não há avaliações
- Fpsyg 11 01662Documento16 páginasFpsyg 11 01662Aldo GuascoAinda não há avaliações
- Functions and Mechanisms of SleepDocumento38 páginasFunctions and Mechanisms of SleepFrancisco MtzAinda não há avaliações
- K-Complexes and Sleep Spindles Before Transient Activation During SleepDocumento8 páginasK-Complexes and Sleep Spindles Before Transient Activation During SleepHernán MarínAinda não há avaliações
- Sleep and Other Interruptions of ConsciousnessDocumento5 páginasSleep and Other Interruptions of ConsciousnessJovelyn NacinAinda não há avaliações
- Normal Sleep and Circadian ProcessesDocumento12 páginasNormal Sleep and Circadian ProcessesHerbert Baquerizo VargasAinda não há avaliações
- Funke Tal 2016Documento9 páginasFunke Tal 2016Luís Eduardo CruvinelAinda não há avaliações
- Saper Et Al. - 2001 - The Sleep Switch Hypothalamic Control of Sleep AnDocumento6 páginasSaper Et Al. - 2001 - The Sleep Switch Hypothalamic Control of Sleep AnNanang HardiansyahAinda não há avaliações
- Dreams Reveal Hidden Meanings and MemoriesDocumento12 páginasDreams Reveal Hidden Meanings and MemoriesNivethithaa DhanrajAinda não há avaliações
- Fre de Ric Bremer 1892-1982 A Pioneer in SleepDocumento10 páginasFre de Ric Bremer 1892-1982 A Pioneer in SleepalejabordaAinda não há avaliações
- Wang 2021Documento9 páginasWang 2021Aldo GuascoAinda não há avaliações
- Neurobiology of The Sleep-Wake Cycle: Sleep Architecture, Circadian Regulation, and Regulatory FeedbackDocumento12 páginasNeurobiology of The Sleep-Wake Cycle: Sleep Architecture, Circadian Regulation, and Regulatory FeedbackRoberto Rafael Ramírez FelipeAinda não há avaliações
- The Neuropsychology of Dreams: A Clinico-Anatomical Study: BooksDocumento2 páginasThe Neuropsychology of Dreams: A Clinico-Anatomical Study: BooksdoraAinda não há avaliações
- Sleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaDocumento12 páginasSleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaMariaAinda não há avaliações
- 1 Introduction PDFDocumento4 páginas1 Introduction PDFMyriam NarváezAinda não há avaliações
- Sleep Medicine 2015 (304 529)Documento226 páginasSleep Medicine 2015 (304 529)Guillermo MoranteAinda não há avaliações
- 123biology of REM SleepDocumento2 páginas123biology of REM SleepJasmin De JesusAinda não há avaliações
- Dreaming and Rem Sleep Are Controlled by DifferDocumento8 páginasDreaming and Rem Sleep Are Controlled by DifferKatja GoiteAinda não há avaliações
- Rapid Eye Movement Rem Sleep Cholinergic Mechanisms1Documento6 páginasRapid Eye Movement Rem Sleep Cholinergic Mechanisms1Matheus VilarinhoAinda não há avaliações
- Sleep-Related Epilepsy Treatment OptionsDocumento17 páginasSleep-Related Epilepsy Treatment OptionsMônica ScattolinAinda não há avaliações
- Induction of Self Awareness in Dreams Through Frontal Low Current Stimulation of Gamma ActivityDocumento5 páginasInduction of Self Awareness in Dreams Through Frontal Low Current Stimulation of Gamma ActivityFelipe CorreacastroAinda não há avaliações
- Makalah Sleep and ConsciounessDocumento4 páginasMakalah Sleep and ConsciounessNabilla Khanza MuzarAinda não há avaliações
- Cerebral Correlates of Delta Waves During non-REM Sleep RevisitedDocumento8 páginasCerebral Correlates of Delta Waves During non-REM Sleep RevisitedEmiliano PrandelliAinda não há avaliações
- Sleep - DMPDocumento53 páginasSleep - DMPdharitri100% (1)
- Sleep ParalysisDocumento7 páginasSleep ParalysisAireeseAinda não há avaliações
- SleepDocumento28 páginasSleepapi-534716659Ainda não há avaliações
- Neural Circuitry of Wakefulness and SleepDocumento40 páginasNeural Circuitry of Wakefulness and Sleepribamar CoutinhoAinda não há avaliações
- Functions and Mechanisms of Sleep: An In-Depth ReviewDocumento36 páginasFunctions and Mechanisms of Sleep: An In-Depth ReviewCintya RambuAinda não há avaliações
- Deep Learning Analysis For Estimating Sleep Syndrome Detection Utilizing The Twin Convolutional Model FTC2Documento7 páginasDeep Learning Analysis For Estimating Sleep Syndrome Detection Utilizing The Twin Convolutional Model FTC2BOHR International Journal of Future Robotics and Artificial Intelligence (BIJFRAI)Ainda não há avaliações
- State of the Art Control of Breathing during SleepDocumento31 páginasState of the Art Control of Breathing during SleepAchmad Hafidz BaraqbahAinda não há avaliações
- Why We Sleep: The Temporal Organization of Recovery: Unsolved MysteryDocumento9 páginasWhy We Sleep: The Temporal Organization of Recovery: Unsolved Mysterystella pangestikaAinda não há avaliações
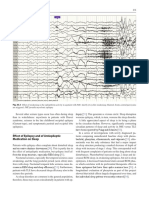
- The Neurologyof Sleep 2012 Published FormDocumento18 páginasThe Neurologyof Sleep 2012 Published FormNeurologie PediatricaAinda não há avaliações
- Lucid Dreaming-A State of Consciousness With Features of Both Waking and Non-Lucid DreamingDocumento10 páginasLucid Dreaming-A State of Consciousness With Features of Both Waking and Non-Lucid DreamingOsvaldo PerezAinda não há avaliações
- Neurobiology of SleepDocumento54 páginasNeurobiology of SleepSwarali 123Ainda não há avaliações
- Vyazovskiy - Sleep and Synaptic Homeostasis - Curr Top Behav Neurosci 2014Documento31 páginasVyazovskiy - Sleep and Synaptic Homeostasis - Curr Top Behav Neurosci 2014Thiago DanieleAinda não há avaliações
- Sleep 02 03Documento21 páginasSleep 02 03Nelu CastravetAinda não há avaliações
- Landmark 1064Documento11 páginasLandmark 1064iyaAinda não há avaliações
- Centennial Review: The Pathogenesis of Obstructive Sleep ApneaDocumento6 páginasCentennial Review: The Pathogenesis of Obstructive Sleep ApneaMelly NoviaAinda não há avaliações
- The Analysis of Brain ActivityDocumento4 páginasThe Analysis of Brain ActivityCabinet VeterinarAinda não há avaliações
- TwoProcessModel PPSChapter BorbleyDocumento16 páginasTwoProcessModel PPSChapter Borbleysuresh2250Ainda não há avaliações
- EEG Normal Sleep - StatPearls - NCBI BookshelfDocumento7 páginasEEG Normal Sleep - StatPearls - NCBI BookshelfChandan ShandilAinda não há avaliações
- NIH Public Access: Control of Sleep and WakefulnessDocumento172 páginasNIH Public Access: Control of Sleep and WakefulnessPrima Akli SetyoAinda não há avaliações
- Alvászavar És MigrénDocumento10 páginasAlvászavar És MigrénLindenberger BoriszAinda não há avaliações
- International Journal of Psychophysiology: Sarah Laxhmi Chellappa, Christian CajochenDocumento6 páginasInternational Journal of Psychophysiology: Sarah Laxhmi Chellappa, Christian CajochenKatja GoiteAinda não há avaliações
- Why We SleepDocumento6 páginasWhy We SleepJustine MenesesAinda não há avaliações
- Yang Dan 2012 (Neuromodulation of Brain States)Documento14 páginasYang Dan 2012 (Neuromodulation of Brain States)FRANCISCO ELI LEZAMA GUTIERREZAinda não há avaliações
- Cote 2002Documento15 páginasCote 2002Chandan ShandilAinda não há avaliações
- CME: Understanding the Science of SleepDocumento4 páginasCME: Understanding the Science of SleepBekir Can ÇelikAinda não há avaliações
- Morales1978 AtoniaDocumento7 páginasMorales1978 AtoniasujAinda não há avaliações
- Dreams, Endocannabinoids and Itinerant Dynamics in Neural NetworksDocumento34 páginasDreams, Endocannabinoids and Itinerant Dynamics in Neural Networksosame kinouchiAinda não há avaliações
- Sleep PresentationDocumento25 páginasSleep PresentationmonicaAinda não há avaliações
- CMB5Documento2 páginasCMB5غسن سمن المدنAinda não há avaliações
- CMB3Documento3 páginasCMB3غسن سمن المدنAinda não há avaliações
- CMB1Documento1 páginaCMB1غسن سمن المدنAinda não há avaliações
- Acanthamoeba In-WPS OfficeDocumento4 páginasAcanthamoeba In-WPS Officeغسن سمن المدنAinda não há avaliações
- Antiplatelet THDocumento3 páginasAntiplatelet THغسن سمن المدنAinda não há avaliações
- CMB2Documento1 páginaCMB2غسن سمن المدنAinda não há avaliações
- CMB4Documento2 páginasCMB4غسن سمن المدنAinda não há avaliações
- Abdominal AorticDocumento6 páginasAbdominal Aorticغسن سمن المدنAinda não há avaliações
- Dan M. Roden-WPS OfficeDocumento1 páginaDan M. Roden-WPS Officeغسن سمن المدنAinda não há avaliações
- AchalasiaDocumento4 páginasAchalasiaغسن سمن المدنAinda não há avaliações
- AbortionDocumento3 páginasAbortionغسن سمن المدنAinda não há avaliações
- Acanthamoeba In-WPS OfficeDocumento4 páginasAcanthamoeba In-WPS Officeغسن سمن المدنAinda não há avaliações
- Recognizing A Pleural EffusionDocumento1 páginaRecognizing A Pleural Effusionغسن سمن المدنAinda não há avaliações
- Lung CollapseDocumento1 páginaLung Collapseغسن سمن المدنAinda não há avaliações
- Emergency Tintin-WPS OfficeDocumento1 páginaEmergency Tintin-WPS Officeغسن سمن المدنAinda não há avaliações
- 17Documento2 páginas17غسن سمن المدنAinda não há avaliações
- Not Seen in This Patient Include:: DiscussionDocumento1 páginaNot Seen in This Patient Include:: Discussionغسن سمن المدنAinda não há avaliações
- Discussion: Pneumonia (Fig. 1a)Documento1 páginaDiscussion: Pneumonia (Fig. 1a)غسن سمن المدنAinda não há avaliações
- Short Incubation PeriodDocumento1 páginaShort Incubation Periodغسن سمن المدنAinda não há avaliações
- Laminar EffusionsDocumento1 páginaLaminar Effusionsغسن سمن المدنAinda não há avaliações
- Types of AtelectasisDocumento1 páginaTypes of Atelectasisغسن سمن المدنAinda não há avaliações
- Imaging of The Breast Is Undertaken As Part of A ComprehensiveDocumento1 páginaImaging of The Breast Is Undertaken As Part of A Comprehensiveغسن سمن المدنAinda não há avaliações
- Recognize Partial or Complete Atelectasis of The Following On A Chest Radiograph or Computed TomographyDocumento1 páginaRecognize Partial or Complete Atelectasis of The Following On A Chest Radiograph or Computed Tomographyغسن سمن المدنAinda não há avaliações
- The Primary Features of Established Illness Are Related To EndothelialDocumento1 páginaThe Primary Features of Established Illness Are Related To Endothelialغسن سمن المدنAinda não há avaliações
- Learning ObjectivesDocumento1 páginaLearning Objectivesغسن سمن المدنAinda não há avaliações
- The Development of The 12Documento1 páginaThe Development of The 12غسن سمن المدنAinda não há avaliações
- Anatomy of Lung Lobes and FissuresDocumento1 páginaAnatomy of Lung Lobes and Fissuresغسن سمن المدنAinda não há avaliações
- Isolated Heart MuscleDocumento1 páginaIsolated Heart Muscleغسن سمن المدنAinda não há avaliações
- A Class I Incisor Relationship Is Defined by The British Standards Incisor Classification As FollowsDocumento2 páginasA Class I Incisor Relationship Is Defined by The British Standards Incisor Classification As Followsغسن سمن المدنAinda não há avaliações
- Anticoagulant andDocumento2 páginasAnticoagulant andغسن سمن المدنAinda não há avaliações
- Epidural and Spinal AnesthesiaDocumento86 páginasEpidural and Spinal Anesthesiasarguss1471% (7)
- Hardy WeinbergDocumento19 páginasHardy Weinbergapi-280302562Ainda não há avaliações
- Photosynthesis Science Presentation in Green Beige Illustrative Style - 20230920 - 192536 - 0000Documento11 páginasPhotosynthesis Science Presentation in Green Beige Illustrative Style - 20230920 - 192536 - 0000Rei B.XAinda não há avaliações
- Two-Dimensional Inverse DynamicsDocumento22 páginasTwo-Dimensional Inverse DynamicsEric Urbina SantibañezAinda não há avaliações
- Connective TissueDocumento57 páginasConnective TissueSunil DhatwaliaAinda não há avaliações
- MBCL Movement ExercisesDocumento5 páginasMBCL Movement ExercisesgustavogknAinda não há avaliações
- KRR 2 1 ElectroDocumento2 páginasKRR 2 1 Electroosiris33ra8248100% (1)
- Cardiogenic Shock HandoutsDocumento2 páginasCardiogenic Shock HandoutsAileenAinda não há avaliações
- Rshakmgp-011r Glp-1 (Active) Elisa Kit Eng.001Documento18 páginasRshakmgp-011r Glp-1 (Active) Elisa Kit Eng.001scribdAinda não há avaliações
- Brain Rules by John Medina (Exercise Chapter)Documento45 páginasBrain Rules by John Medina (Exercise Chapter)Pear Press100% (6)
- Spleen (Chinese Medicine)Documento2 páginasSpleen (Chinese Medicine)Doktormin106Ainda não há avaliações
- PNLE Sample Test QuestionsDocumento12 páginasPNLE Sample Test QuestionsHam-Ham LootAinda não há avaliações
- Abigail Domingo Physical AssessmentDocumento87 páginasAbigail Domingo Physical AssessmentVenus Maui RoqueAinda não há avaliações
- J. Biol. Chem.-1989-Boggaram-11421-7Documento7 páginasJ. Biol. Chem.-1989-Boggaram-11421-7Patrick RamosAinda não há avaliações
- Calcarea fluorica remedy for fear of instability and povertyDocumento16 páginasCalcarea fluorica remedy for fear of instability and povertycgarciAinda não há avaliações
- The Fundamental Unit of Life To Improvement in Food Resources ChaptersDocumento104 páginasThe Fundamental Unit of Life To Improvement in Food Resources ChaptersSARANSH oPAinda não há avaliações
- Author 'S Accepted ManuscriptDocumento46 páginasAuthor 'S Accepted ManuscriptRaul DoctoAinda não há avaliações
- Cambridge IGCSE: Biology 0610/22Documento20 páginasCambridge IGCSE: Biology 0610/22Defaults rulezAinda não há avaliações
- Branford High SchoolDocumento1 páginaBranford High SchoolHelen BennettAinda não há avaliações
- VITAL SIGNS ReviewDocumento4 páginasVITAL SIGNS ReviewA CAinda não há avaliações
- Main Sulci & Fissures: Cerebral FissureDocumento17 páginasMain Sulci & Fissures: Cerebral FissureNagbhushan BmAinda não há avaliações
- Monosaccharides: AlpineDocumento12 páginasMonosaccharides: AlpineReigner Jay B. EscartinAinda não há avaliações
- Immune Dysregulation Among Students Exposed To Exam Stress and Its Mitigation by Mindfulness Training - Findings From An Exploratory Randomised TrialDocumento11 páginasImmune Dysregulation Among Students Exposed To Exam Stress and Its Mitigation by Mindfulness Training - Findings From An Exploratory Randomised TrialPatricia Elena ManaliliAinda não há avaliações
- Chapter 12 Key TermsDocumento3 páginasChapter 12 Key TermsAlthea MangasarAinda não há avaliações
- Aristotle's History of Animals Book IDocumento474 páginasAristotle's History of Animals Book IsenadbeyAinda não há avaliações
- Animal Tissues and Organ SystemsDocumento47 páginasAnimal Tissues and Organ SystemsBenjamin Fernandez Jr.Ainda não há avaliações
- Fermented Milk Products May Improve HealthDocumento7 páginasFermented Milk Products May Improve Healthmilu1312Ainda não há avaliações
- Conceito de Pré Carga e Pós Carga em CardiologiaDocumento9 páginasConceito de Pré Carga e Pós Carga em CardiologiarmesquitajrAinda não há avaliações
- Smooth & Cardiac Muscles: DR Marlini Muhamad FPSK, Usim Nov 2012Documento34 páginasSmooth & Cardiac Muscles: DR Marlini Muhamad FPSK, Usim Nov 2012Harleyquinn96 DrAinda não há avaliações
- Acid-Base Tutorial - Kerry BrandisDocumento159 páginasAcid-Base Tutorial - Kerry BrandisJason TiongAinda não há avaliações