Você também pode gostar
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (121)
- Region 2 - Community-Based Services - September 2017Documento12 páginasRegion 2 - Community-Based Services - September 2017Jennifer BankerAinda não há avaliações
- First Aid Cdrrmo 2019Documento373 páginasFirst Aid Cdrrmo 2019fenan sollanoAinda não há avaliações
- Malaprabha Biogas Plant Developed by DR S V MapuskarDocumento40 páginasMalaprabha Biogas Plant Developed by DR S V Mapuskarsvmapuskar4914100% (3)
- Case Study Sa Surgical WardDocumento8 páginasCase Study Sa Surgical WardAiza ToledanaAinda não há avaliações
- NCP and CNPDocumento37 páginasNCP and CNPDen TupasAinda não há avaliações
- Journal of Clinical Gerontology & Geriatrics: Camilla Jing Hwa Chern, BS, Shyh-Dye Lee, MD, MPHDocumento5 páginasJournal of Clinical Gerontology & Geriatrics: Camilla Jing Hwa Chern, BS, Shyh-Dye Lee, MD, MPHEdith Frederick LiemAinda não há avaliações
- CHECKLISTDocumento7 páginasCHECKLISTMichelle CardeñoAinda não há avaliações
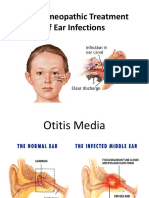
- The Homeopathic Treatment of Ear Infections PDFDocumento104 páginasThe Homeopathic Treatment of Ear Infections PDFdipgang7174Ainda não há avaliações
- Abdominal Aortic AneurysmDocumento7 páginasAbdominal Aortic AneurysmShaastieAinda não há avaliações
- Chapter 102: Laryngeal Trauma From Intubation: Endoscopic Evaluation and Classification Bruce BenjaminDocumento16 páginasChapter 102: Laryngeal Trauma From Intubation: Endoscopic Evaluation and Classification Bruce BenjaminPrisia AnantamaAinda não há avaliações
- Contoh IjapDocumento4 páginasContoh IjapFarihaAinda não há avaliações
- Water Quality at Residential and Industrial AreaDocumento11 páginasWater Quality at Residential and Industrial AreaMohamad AizanAinda não há avaliações
- Leptospirosis CaseDocumento29 páginasLeptospirosis CaseJp RectraAinda não há avaliações
- Essential Fatty Acid Flax EPA ALADocumento7 páginasEssential Fatty Acid Flax EPA ALAlakshmibava100% (1)
- Health Benefits of HoneyDocumento10 páginasHealth Benefits of HoneyMohd Idris MohiuddinAinda não há avaliações
- Ocular Manifestations in AIDSDocumento27 páginasOcular Manifestations in AIDSAnumeha Jindal100% (1)
- Comisión LancetDocumento66 páginasComisión LancetKaya CromoAinda não há avaliações
- Glossary of Traditional Chinese Medicine 中醫術語Documento7 páginasGlossary of Traditional Chinese Medicine 中醫術語OscarTadlockAinda não há avaliações
- Patient Profile Data Form: Advance DirectivesDocumento6 páginasPatient Profile Data Form: Advance DirectivesMa Cecilia SorianoAinda não há avaliações
- Wallen Et Al-2006-Australian Occupational Therapy JournalDocumento1 páginaWallen Et Al-2006-Australian Occupational Therapy Journal胡知行Ainda não há avaliações
- Movement Diagrams - Rayner & Smale PDFDocumento6 páginasMovement Diagrams - Rayner & Smale PDFItai IzhakAinda não há avaliações
- Informed Consent Form Whole30 2017Documento1 páginaInformed Consent Form Whole30 2017api-385355641Ainda não há avaliações
- Mera MerckDocumento5 páginasMera Merckkuch bhi100% (1)
- Samantha Nord-Shafer vs. Narconon: ComplaintDocumento33 páginasSamantha Nord-Shafer vs. Narconon: ComplaintTony OrtegaAinda não há avaliações
- Lesson 6 Documentation of Pharmacotherapy InterventionsDocumento1 páginaLesson 6 Documentation of Pharmacotherapy InterventionsVincent Joshua TriboAinda não há avaliações
- Imp Questions in PharmacologyDocumento5 páginasImp Questions in PharmacologyBeerasandrapalyaRangaiahRavikumar100% (3)
- Lab 8 - Self Mon3Documento48 páginasLab 8 - Self Mon3pinkCherri100% (4)
- Aerosol TherapyDocumento10 páginasAerosol Therapy04lubna_869632400Ainda não há avaliações
- IndianJPain3341-1062069 025700Documento10 páginasIndianJPain3341-1062069 025700docbinAinda não há avaliações
- LUTRONIC 2020 CataolgDocumento13 páginasLUTRONIC 2020 Cataolgkhairul ihsanAinda não há avaliações