Você também pode gostar
- Lab 5Documento2 páginasLab 5Cory MancusoAinda não há avaliações
- Pharm Chem HWDocumento4 páginasPharm Chem HWCory MancusoAinda não há avaliações
- Slaughterhouse Five As A Postmodernist WorkDocumento2 páginasSlaughterhouse Five As A Postmodernist WorkCory MancusoAinda não há avaliações
- Alien II GB 0911Documento2 páginasAlien II GB 0911Cory MancusoAinda não há avaliações
- Slaughterhouse Five As A Postmodernist WorkDocumento2 páginasSlaughterhouse Five As A Postmodernist WorkCory MancusoAinda não há avaliações
- SomethingDocumento6 páginasSomethingCory MancusoAinda não há avaliações
- Chap2b MDocumento39 páginasChap2b MCory MancusoAinda não há avaliações
- 220 - Lecture 1 - D2LDocumento32 páginas220 - Lecture 1 - D2LCory MancusoAinda não há avaliações
- Research PaperDocumento7 páginasResearch PaperCory MancusoAinda não há avaliações
- # Title Director Ratin G ReviewDocumento5 páginas# Title Director Ratin G ReviewCory MancusoAinda não há avaliações
- Advance Directives & Surrogate Decision-Makers: Rights & ResponsibilitiesDocumento24 páginasAdvance Directives & Surrogate Decision-Makers: Rights & ResponsibilitiesCory MancusoAinda não há avaliações
- # Title Director Ratin G ReviewDocumento5 páginas# Title Director Ratin G ReviewCory MancusoAinda não há avaliações
- MoviesDocumento1 páginaMoviesCory MancusoAinda não há avaliações
- Carly Comando EverydayDocumento7 páginasCarly Comando Everydaycezzyd60% (5)
- ApricotsDocumento3 páginasApricotsCory MancusoAinda não há avaliações
- ApricotsDocumento3 páginasApricotsCory MancusoAinda não há avaliações
- Movies To SeeDocumento2 páginasMovies To SeeCory MancusoAinda não há avaliações
- Castlevania 2 Bloody Tears Sheet MusicDocumento1 páginaCastlevania 2 Bloody Tears Sheet MusicReda LahlouAinda não há avaliações
- Lonely Man IIDocumento11 páginasLonely Man IIBill RillAinda não há avaliações
- Russian NounsDocumento3 páginasRussian NounsCory MancusoAinda não há avaliações
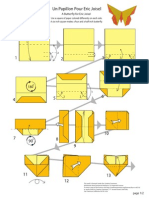
- Papillon PDFDocumento2 páginasPapillon PDFJuan Sebastian Nieto PreciadoAinda não há avaliações
- Atomic Structure and Quantum ModelsDocumento2 páginasAtomic Structure and Quantum ModelsCory MancusoAinda não há avaliações
- Calculus AnswersDocumento560 páginasCalculus AnswersCory Mancuso50% (2)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNo EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeNota: 4 de 5 estrelas4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingNo EverandThe Little Book of Hygge: Danish Secrets to Happy LivingNota: 3.5 de 5 estrelas3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNo EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceNota: 4 de 5 estrelas4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNo EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureNota: 4.5 de 5 estrelas4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)No EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Nota: 4 de 5 estrelas4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnNo EverandTeam of Rivals: The Political Genius of Abraham LincolnNota: 4.5 de 5 estrelas4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItNo EverandNever Split the Difference: Negotiating As If Your Life Depended On ItNota: 4.5 de 5 estrelas4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerNo EverandThe Emperor of All Maladies: A Biography of CancerNota: 4.5 de 5 estrelas4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNo EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryNota: 3.5 de 5 estrelas3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNo EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaNota: 4.5 de 5 estrelas4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNo EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersNota: 4.5 de 5 estrelas4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyNo EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyNota: 3.5 de 5 estrelas3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaNo EverandThe Unwinding: An Inner History of the New AmericaNota: 4 de 5 estrelas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNo EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreNota: 4 de 5 estrelas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)No EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Nota: 4.5 de 5 estrelas4.5/5 (119)
- Final CirculatoryDocumento44 páginasFinal CirculatoryKeiko-Kaori NebresAinda não há avaliações
- Unit D - Human Systems Unit PlanDocumento4 páginasUnit D - Human Systems Unit Planapi-427321002Ainda não há avaliações
- Achieving Excellence in The OSCE Part 1Documento409 páginasAchieving Excellence in The OSCE Part 1Sultan AlexandruAinda não há avaliações
- B.inggris KtiDocumento61 páginasB.inggris KtiDimasAinda não há avaliações
- Manage Cardiac Output in Hyperthyroid PatientDocumento3 páginasManage Cardiac Output in Hyperthyroid PatientPrincess QuirinaAinda não há avaliações
- Natalya Master CardioDocumento71 páginasNatalya Master CardioTaman Hoang100% (1)
- Concept Strengthening Sheet CSS-03 Zoology: Based On AIATS-03 TYMDocumento3 páginasConcept Strengthening Sheet CSS-03 Zoology: Based On AIATS-03 TYMSparsh MehtaAinda não há avaliações
- Cardiovascular System Histo LabDocumento53 páginasCardiovascular System Histo Labsultan khabeebAinda não há avaliações
- Cardiac Catheterization ProcedureDocumento10 páginasCardiac Catheterization ProcedureDimpal Choudhary100% (4)
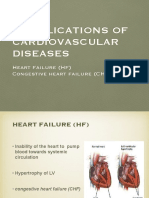
- V. Complications of CVDDocumento26 páginasV. Complications of CVDJan Federick BantayAinda não há avaliações
- Life Processes Part III: Transportation in Humans and PlantsDocumento4 páginasLife Processes Part III: Transportation in Humans and PlantsApoorv ChauhanAinda não há avaliações
- 06 Heart SoundsDocumento24 páginas06 Heart SoundsDolphinAinda não há avaliações
- J Jacc 2023 01 049 PDFDocumento25 páginasJ Jacc 2023 01 049 PDFRaquel AguilarAinda não há avaliações
- Sistem SirkulasiDocumento21 páginasSistem SirkulasiRisna YusniAinda não há avaliações
- Angina Pectoris PathophysiologyDocumento4 páginasAngina Pectoris Pathophysiologyteddydeclines1467% (6)
- Development, Basic Goals and Concepts in MyocardialDocumento13 páginasDevelopment, Basic Goals and Concepts in MyocardialAtia KiranAinda não há avaliações
- Manual of Pediatric Cardiac Intensive CareDocumento326 páginasManual of Pediatric Cardiac Intensive CareLeydi Baltazar T100% (4)
- B7.2 Transport in MammalsDocumento13 páginasB7.2 Transport in Mammalsulavan24Ainda não há avaliações
- Nursing Care Plan For Palliative Care Nursing NotesDocumento2 páginasNursing Care Plan For Palliative Care Nursing NotesblaireAinda não há avaliações
- Cardio TransDocumento7 páginasCardio TransweissAinda não há avaliações
- B.pharmacy Part-I (Sem I & II)Documento37 páginasB.pharmacy Part-I (Sem I & II)gocrazy with yashAinda não há avaliações
- Acyanotic Disease: Classification of CHDDocumento11 páginasAcyanotic Disease: Classification of CHDFenita Renny DinataAinda não há avaliações
- Sikkim University: Rules & Syllabus For The Bachelor of Pharmacy (B. Pharm) CourseDocumento147 páginasSikkim University: Rules & Syllabus For The Bachelor of Pharmacy (B. Pharm) CourseVikash KushwahaAinda não há avaliações
- ACCUVIX XG Reference Manual E PDFDocumento274 páginasACCUVIX XG Reference Manual E PDFMonica FajardoAinda não há avaliações
- Pulmonary TraumatologyDocumento15 páginasPulmonary Traumatologynoor88Ainda não há avaliações
- Pathology Slides 1Documento28 páginasPathology Slides 1SydAinda não há avaliações
- Pathophysiology and Etiology of Edema in AdultsDocumento15 páginasPathophysiology and Etiology of Edema in AdultsJovie Anne CabangalAinda não há avaliações
- Heart Dissection WorksheetDocumento5 páginasHeart Dissection Worksheetapi-237875296100% (1)
- ANP 1105 The Lymphatic SystemDocumento23 páginasANP 1105 The Lymphatic SystemMathios TigerosAinda não há avaliações
- SVT PresentationDocumento39 páginasSVT PresentationReinsy NoviAinda não há avaliações