Você também pode gostar
- A. Homeostasis Autocontrol BiofisicaDocumento45 páginasA. Homeostasis Autocontrol BiofisicaJavier AcuñaAinda não há avaliações
- Agosto 3, 2009 Lunes S - Ntesis de Prote - NasDocumento12 páginasAgosto 3, 2009 Lunes S - Ntesis de Prote - NasJavier AcuñaAinda não há avaliações
- Agricultura AsociativaDocumento7 páginasAgricultura AsociativaJavier AcuñaAinda não há avaliações
- Las Membranas Biológicas y TransporteDocumento33 páginasLas Membranas Biológicas y TransporteJavier AcuñaAinda não há avaliações
- 3 INFORME Producciones Ovina y BovinaDocumento11 páginas3 INFORME Producciones Ovina y BovinaJavier AcuñaAinda não há avaliações
- La Organización de Las CélulasDocumento15 páginasLa Organización de Las CélulasJavier AcuñaAinda não há avaliações
- Huevos de Gallina CriollosDocumento3 páginasHuevos de Gallina CriollosJavier AcuñaAinda não há avaliações
- GallinasDocumento61 páginasGallinasSergio Andres Aparicio GalvisAinda não há avaliações
- B8 A4 FD 01Documento62 páginasB8 A4 FD 01Marco Saez MurilloAinda não há avaliações
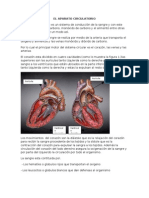
- El Aparato CirculatorioDocumento2 páginasEl Aparato CirculatorioJavier AcuñaAinda não há avaliações
- Guia 1 - Introduccion - pdfeXCELDocumento12 páginasGuia 1 - Introduccion - pdfeXCELSerena TaylorAinda não há avaliações
- Histologia Arterias Venas y CapilaresDocumento45 páginasHistologia Arterias Venas y CapilaresJorge Bautistad100% (3)
- Documento Teórico ... Ciclo de KrebsDocumento6 páginasDocumento Teórico ... Ciclo de KrebsandreinaAinda não há avaliações
- P 1 Presion USMP 2021Documento80 páginasP 1 Presion USMP 2021Erika Ruiz TorresAinda não há avaliações
- TAREA 02 SIST - CIRCULATORIO Sem 02Documento2 páginasTAREA 02 SIST - CIRCULATORIO Sem 02Milagros HidalgoAinda não há avaliações
- Ciclo KerbsDocumento3 páginasCiclo KerbsAllan Kun DamasAinda não há avaliações
- Actividad 1 y 2 de La Semana 12 - Gianfranco Talavera GutierrezDocumento5 páginasActividad 1 y 2 de La Semana 12 - Gianfranco Talavera GutierrezGIANFRANCO TALAVERA GUTIERREZAinda não há avaliações
- C - S - I - Son Nesesarias Las Plantas para VivirDocumento5 páginasC - S - I - Son Nesesarias Las Plantas para VivirFabiola Mena EspinozaAinda não há avaliações
- Cuadro ComparativoDocumento3 páginasCuadro ComparativoAstrid Ramirez OrozcoAinda não há avaliações
- Sistema CirculatorioDocumento71 páginasSistema CirculatorioNilLeninPalaciosCamacho100% (1)
- Sistema CirculatorioDocumento2 páginasSistema CirculatorioChristian LinaresAinda não há avaliações
- Cadena Transportadora de ElectronesDocumento5 páginasCadena Transportadora de ElectronesAmadaGarciaYepezAinda não há avaliações
- PRÁCTICA No 6. 06 - 2023 Procesos RespiratoriosDocumento17 páginasPRÁCTICA No 6. 06 - 2023 Procesos RespiratorioslpalacioscaAinda não há avaliações
- Mitocondrias y Respiracion CelualrDocumento31 páginasMitocondrias y Respiracion Celualrwilliams rolando huaraca pomaAinda não há avaliações
- Tarea BenryDocumento3 páginasTarea Benryhenry Miranda ArteagaAinda não há avaliações
- Ac17 MedulaDocumento18 páginasAc17 Medulajuan peres peresAinda não há avaliações
- Tema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Documento15 páginasTema 15 Cadena Respiratoria. Autor Alexander Sanchez Lamas.Alexander SanchezAinda não há avaliações
- Circulacion MenorDocumento1 páginaCirculacion MenorYeny LermaAinda não há avaliações
- Informe de Laboratorio 7Documento6 páginasInforme de Laboratorio 7SthefiiEnriquezAinda não há avaliações
- Informe de Laboratorio N°9Documento7 páginasInforme de Laboratorio N°9JuniorAinda não há avaliações
- 4 Biotransformacion de La EnergíaDocumento4 páginas4 Biotransformacion de La EnergíaErick Keyser Abanto VillarrealAinda não há avaliações
- Anatomia y Fisiologia Del CorazonDocumento26 páginasAnatomia y Fisiologia Del CorazonJonathan Andres Franco SánchezAinda não há avaliações
- Taller, BiologíaDocumento18 páginasTaller, BiologíaAdrian CorredorAinda não há avaliações
- Ciclo CardiacoDocumento3 páginasCiclo CardiacoFranliz ZambranoAinda não há avaliações
- Cadena Transportadora de ElectronesDocumento12 páginasCadena Transportadora de ElectronesReinaldo Bajo100% (1)
- Tema 17. CatabolismoDocumento80 páginasTema 17. CatabolismoPepito GrilloAinda não há avaliações
- 6 Sistema de Conducción Del CorazónDocumento2 páginas6 Sistema de Conducción Del CorazónMildred YamilethAinda não há avaliações
- Angela Herrera MDocumento6 páginasAngela Herrera MYubraska Marquez HurtadoAinda não há avaliações
- Continuum Energetico en NatacionDocumento22 páginasContinuum Energetico en NatacionMiguel Padilla100% (2)
- Aparato Cardiovascular para Cuarto de Primaria PDFDocumento4 páginasAparato Cardiovascular para Cuarto de Primaria PDFCindy Correa Rangel100% (1)
- Gluconeogenesis PDFDocumento26 páginasGluconeogenesis PDFVerónica PovedaAinda não há avaliações