Você também pode gostar
- Fundamentos y técnicas básicas en biología molecularNo EverandFundamentos y técnicas básicas en biología molecularAinda não há avaliações
- Nucleo ArquitecturaDocumento8 páginasNucleo Arquitecturaarcadia_hbAinda não há avaliações
- Los límites entre la histología y la bioquímica: observando al núcleo celular con ojo histológico-bioquímicoDocumento16 páginasLos límites entre la histología y la bioquímica: observando al núcleo celular con ojo histológico-bioquímicoedson floresAinda não há avaliações
- Reporte Del Nucleo CelularDocumento4 páginasReporte Del Nucleo Celularacademia puruaran100% (1)
- Articulos CientificosDocumento4 páginasArticulos CientificosAlejandra Arcos GuerreroAinda não há avaliações
- Histología, Embriología, Biología Molecular y GenéticaDocumento3 páginasHistología, Embriología, Biología Molecular y GenéticaElisa LaraAinda não há avaliações
- Guía TP 5Documento16 páginasGuía TP 5Dani Morales DellatorreAinda não há avaliações
- La Frontera Genetica Molecular de La EnfermedadDocumento526 páginasLa Frontera Genetica Molecular de La EnfermedadDeivy Horta Tkd0% (1)
- Células del sistema nerviosoDocumento12 páginasCélulas del sistema nerviosoSamayraAinda não há avaliações
- Envuelta nuclear, el protector del núcleoDocumento20 páginasEnvuelta nuclear, el protector del núcleomanuel rodriguesAinda não há avaliações
- Práctica 3..Documento4 páginasPráctica 3..hannacastroortizAinda não há avaliações
- Núcleo Celular - Wikipedia, La Enciclopedia LibreDocumento134 páginasNúcleo Celular - Wikipedia, La Enciclopedia LibreruizyannarithAinda não há avaliações
- Cuadernillo de Biologia 1 Bloque Iii - ZambranoDocumento23 páginasCuadernillo de Biologia 1 Bloque Iii - ZambranoMarcos Zambrano AlegriaAinda não há avaliações
- Trabajo Práctico #2.Documento23 páginasTrabajo Práctico #2.Juan Cruz FARFANAinda não há avaliações
- Ensayo-El Nucleo CelularDocumento18 páginasEnsayo-El Nucleo CelularDarwin TapuyoAinda não há avaliações
- Genética SeminarioDocumento7 páginasGenética SeminarioMayerling V. RiosAinda não há avaliações
- Sistema Nervioso y Vesiculas CerebralesDocumento35 páginasSistema Nervioso y Vesiculas Cerebraleslorietarosemary100% (1)
- La Frontera Genetica - MolecularDocumento526 páginasLa Frontera Genetica - MolecularFrancis Vallecillo PadillaAinda não há avaliações
- PPT-Sesión 08Documento20 páginasPPT-Sesión 08Andre100% (1)
- Nucleo InterfasticoDocumento18 páginasNucleo InterfasticoAnahi Alban100% (1)
- Tema 10_El núcleo_Reproducción celularDocumento23 páginasTema 10_El núcleo_Reproducción celularcandelaropero2006Ainda não há avaliações
- Unidad 09 ReplicacionDocumento12 páginasUnidad 09 ReplicacionMicaela FragaAinda não há avaliações
- Unidad 9 ReplicDocumento29 páginasUnidad 9 ReplicLUDMILA FERNÁNDEZAinda não há avaliações
- El Nucléolo Como Un Regulador Del Envejecimiento CelularDocumento19 páginasEl Nucléolo Como Un Regulador Del Envejecimiento CelularRandolfo MaximilianoAinda não há avaliações
- Cell Mechanics and The CytoskeletonDocumento22 páginasCell Mechanics and The CytoskeletonJohn Fredy Moreno MolanoAinda não há avaliações
- Núcleo Celular y MitosisDocumento8 páginasNúcleo Celular y MitosisNathalia GonzalezAinda não há avaliações
- El núcleo celular: estructura y funciones claveDocumento20 páginasEl núcleo celular: estructura y funciones claveCarina BugliaAinda não há avaliações
- Primera Actividad Biologia UdecDocumento1 páginaPrimera Actividad Biologia UdecAngie RodriguezAinda não há avaliações
- Organelas Celulares: Núcleo y Ácidos NucleicosDocumento17 páginasOrganelas Celulares: Núcleo y Ácidos NucleicosMarcela BarriosAinda não há avaliações
- Artículo Científico - La Biología Molecular y El ADNDocumento6 páginasArtículo Científico - La Biología Molecular y El ADNJosé PaterninaAinda não há avaliações
- mdml502 s1 Valenzuela1Documento10 páginasmdml502 s1 Valenzuela1Larissa Noemi Saavedra ZuberAinda não há avaliações
- Clase Estructura Del NucleoDocumento90 páginasClase Estructura Del Nucleohghbkgykmgxyh100% (1)
- Resumen Febrero 2p.okDocumento10 páginasResumen Febrero 2p.okMaría JoséAinda não há avaliações
- Teoría CelularDocumento4 páginasTeoría Celularto son círculosAinda não há avaliações
- Texto Ilustrado de Biologia Molecular e Ingenieria Genetica 2da Edicion - Angel Herraez PDFDocumento504 páginasTexto Ilustrado de Biologia Molecular e Ingenieria Genetica 2da Edicion - Angel Herraez PDFJulio Gonzalez Vasquez100% (3)
- Biología MolecularDocumento4 páginasBiología MolecularIrandini MonroyAinda não há avaliações
- Laboratorio No. 2Documento2 páginasLaboratorio No. 2EnmanuelAinda não há avaliações
- Informe de Exposición NúcleoDocumento14 páginasInforme de Exposición Núcleomafer salazarAinda não há avaliações
- Guia de Estudio - NucleoDocumento3 páginasGuia de Estudio - NucleoAzul NordenstromAinda não há avaliações
- Universidad Nacional Pedro Ruiz GalloDocumento22 páginasUniversidad Nacional Pedro Ruiz GalloBrain Jonatan SullonAinda não há avaliações
- El Nucleo CelularDocumento13 páginasEl Nucleo CelularMauricio AmtAinda não há avaliações
- Nucleo en BiologiaDocumento15 páginasNucleo en BiologiaAnnie BalderasAinda não há avaliações
- Unidad 1 - Tema 2 - CelulasDocumento47 páginasUnidad 1 - Tema 2 - CelulasDai TabordaAinda não há avaliações
- FROTISDocumento5 páginasFROTISElqui MeraAinda não há avaliações
- Teoría CelularDocumento4 páginasTeoría Celularwilfrido herreraAinda não há avaliações
- Estructura Y Función Celular - Loewy Y SiekevitzDocumento233 páginasEstructura Y Función Celular - Loewy Y SiekevitzFabrizzio Piffaretti Romero100% (1)
- Fichero de CelulaDocumento26 páginasFichero de CelulaEMMANUEL SANCHEZ0% (1)
- El núcleo celular: estructura y funciones principalesDocumento17 páginasEl núcleo celular: estructura y funciones principalesVannia Rodriguez MartinezAinda não há avaliações
- Ultraestructura de MembranaDocumento4 páginasUltraestructura de MembranaDIANA CAROLINA PEREZ ACOSTAAinda não há avaliações
- Cell_Mechanics_and_the_Cytoskeleton (1) traducciónDocumento22 páginasCell_Mechanics_and_the_Cytoskeleton (1) traducciónManuela Julieth Barrera LópezAinda não há avaliações
- TEMA Citologia IV (Nucleo de La Celula) (2022 - 2023)Documento10 páginasTEMA Citologia IV (Nucleo de La Celula) (2022 - 2023)pairumanikellyAinda não há avaliações
- Guía Biología C - Octubre 2016 - Versión ColorDocumento139 páginasGuía Biología C - Octubre 2016 - Versión ColorBel Minda CostaAinda não há avaliações
- Guía Biología C - Octubre 2016 - Versión Color PDFDocumento139 páginasGuía Biología C - Octubre 2016 - Versión Color PDFAlvaro Diego CastilloAinda não há avaliações
- Nucleo CelularDocumento8 páginasNucleo CelularJHARIM NOELY AUCCAPIÑA GUILLENAinda não há avaliações
- Ravassi AspectosDocumento33 páginasRavassi AspectosJhon RVAinda não há avaliações
- Insuf Renal Aguda RIFLEDocumento17 páginasInsuf Renal Aguda RIFLEJuan RamirezAinda não há avaliações
- Prolapso Genital Femenino - Lo Que Deberia SaberDocumento8 páginasProlapso Genital Femenino - Lo Que Deberia SaberJhon RVAinda não há avaliações
- Distres RespiratorioDocumento21 páginasDistres RespiratorioBelissa Denisse Calbay CobaAinda não há avaliações
- Análisis psicométrico de la prueba BIAS en estudiantesDocumento19 páginasAnálisis psicométrico de la prueba BIAS en estudiantesJhon RVAinda não há avaliações
- 1 Origen de La Vidamundo ArnDocumento15 páginas1 Origen de La Vidamundo ArnJhon RVAinda não há avaliações
- Crisis AsmaticaDocumento11 páginasCrisis AsmaticaJhon RVAinda não há avaliações
- HipoglucemiaDocumento15 páginasHipoglucemiaJhon RVAinda não há avaliações
- Protocolo Estatus Asmático 2013Documento25 páginasProtocolo Estatus Asmático 2013Anayeli VaFanAinda não há avaliações
- Curb 65Documento1 páginaCurb 65Tuto Vera-BrionesAinda não há avaliações
- Protocolo Diagnóstico y Terapéutico PDFDocumento5 páginasProtocolo Diagnóstico y Terapéutico PDFJhon RVAinda não há avaliações
- Neumonia NosocomialDocumento6 páginasNeumonia NosocomialJhon RVAinda não há avaliações
- Norma Tecnica 106 Recien Nacido PDFDocumento31 páginasNorma Tecnica 106 Recien Nacido PDFDaniel Gavidia0% (1)
- Bioseguridad MedicaDocumento66 páginasBioseguridad MedicaodjhoAinda não há avaliações
- Pneumocystis Jiroveci (Carinii) (3F6) MEN ES IVD 0.0Documento6 páginasPneumocystis Jiroveci (Carinii) (3F6) MEN ES IVD 0.0Jhon RVAinda não há avaliações
- Edema Bene Espanola UciDocumento3 páginasEdema Bene Espanola UciJamesonQPAinda não há avaliações
- Metabolismo Del GlucógenoDocumento6 páginasMetabolismo Del GlucógenoJhon RVAinda não há avaliações
- FX de La Orbita y LefortDocumento23 páginasFX de La Orbita y LefortKatherine Ramirez QuispeAinda não há avaliações
- Glucolisis PDFDocumento3 páginasGlucolisis PDFEstefania Arizaga100% (1)
- Purificación proteínas 6 pasosDocumento8 páginasPurificación proteínas 6 pasosBri Gonzalez TrejoAinda não há avaliações
- Curb 65Documento1 páginaCurb 65Tuto Vera-BrionesAinda não há avaliações
- Coagulacion 2009 Una Vision Moderna de La HemostasiaDocumento5 páginasCoagulacion 2009 Una Vision Moderna de La HemostasiaArturo Canseco LopezAinda não há avaliações
- A 10 V 56 N 1Documento13 páginasA 10 V 56 N 1Ramos LuketyAinda não há avaliações
- Secrecion de ProlactinaDocumento6 páginasSecrecion de ProlactinaDiana BOAinda não há avaliações
- Caso 2Documento3 páginasCaso 2Marisol JaramilloAinda não há avaliações
- Dolor Abdominal. AgudoDocumento9 páginasDolor Abdominal. AgudoJhon RVAinda não há avaliações
- Alteraciones Ungueales Como Marcador de Enfermedad SistémicaDocumento7 páginasAlteraciones Ungueales Como Marcador de Enfermedad SistémicamerodeadorzAinda não há avaliações
- Epilepsia JAMADocumento1 páginaEpilepsia JAMADiana BOAinda não há avaliações
- Interpretación Del HemogramaDocumento15 páginasInterpretación Del HemogramaBárbara Yáñez OsorioAinda não há avaliações
- Taller de BiologíaDocumento10 páginasTaller de BiologíavalAinda não há avaliações
- 12 Biolog°aDocumento8 páginas12 Biolog°aEliezer AbregoAinda não há avaliações
- PreguntaDocumento5 páginasPreguntaJG SanchezAinda não há avaliações
- La Necrofauna y La ModernidadDocumento4 páginasLa Necrofauna y La ModernidadOsvaldoAinda não há avaliações
- Bases Genéticas de Las EnfermedadesDocumento9 páginasBases Genéticas de Las EnfermedadesRomyna Andrea MonsalvesAinda não há avaliações
- Guía N°2 - Ciclo Celular - MITOSIS - II-medioDocumento9 páginasGuía N°2 - Ciclo Celular - MITOSIS - II-medioregulus blackAinda não há avaliações
- Garcia Moro Ruth PFM2Documento70 páginasGarcia Moro Ruth PFM2lugh88Ainda não há avaliações
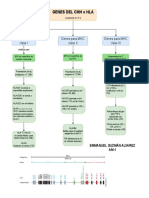
- Genes de HLA o MHCDocumento1 páginaGenes de HLA o MHCEmmanuel GAinda não há avaliações
- Cuestionario Prueba 2 CoreDocumento8 páginasCuestionario Prueba 2 CoreJorge VerdejoAinda não há avaliações
- Caracteres Del Guisante Analizados Por MendelDocumento2 páginasCaracteres Del Guisante Analizados Por MendelgtabimaelAinda não há avaliações
- Clonación de ADNDocumento21 páginasClonación de ADNbryan albanAinda não há avaliações
- Resumen "Fundamentos de Genetica"Documento2 páginasResumen "Fundamentos de Genetica"Kristen Azbelth Coronel MunozAinda não há avaliações
- Seminario 2 Biología Molecular Replicación Adn 2021 Subgrupo2 m622Documento14 páginasSeminario 2 Biología Molecular Replicación Adn 2021 Subgrupo2 m622Erick M. Rivera A.Ainda não há avaliações
- Cromosomas Sexuales y AutosomasDocumento2 páginasCromosomas Sexuales y AutosomasSara Elizabeth Flores70% (10)
- Problemas Morales de La Práctica Médica y La BioéticaDocumento44 páginasProblemas Morales de La Práctica Médica y La BioéticaAbi AlonsoAinda não há avaliações
- Actividades para Modelar: Nombre: Grado: FechaDocumento1 páginaActividades para Modelar: Nombre: Grado: FechaBreicen Andrea Acevedo Vargas Acevedo VargasAinda não há avaliações
- Apuntes Bioquimica Udemy Maestro Camach Leanr Ingeniero en BiotecnologíaDocumento2 páginasApuntes Bioquimica Udemy Maestro Camach Leanr Ingeniero en BiotecnologíaHader CruzAinda não há avaliações
- Que Son Las MutacionesDocumento3 páginasQue Son Las Mutacionesgissell26328174Ainda não há avaliações
- Trab. Biofisica DaltonismoDocumento15 páginasTrab. Biofisica DaltonismoMatheus Felipe100% (1)
- Cariotipo y Mutaciones BiologiaDocumento8 páginasCariotipo y Mutaciones BiologiaLiliana ElizabethAinda não há avaliações
- Evaluación Diagnóstica: Nombre: Grado: FechaDocumento2 páginasEvaluación Diagnóstica: Nombre: Grado: FechaAna MatamorosAinda não há avaliações
- MutacionesDocumento11 páginasMutacionesPaola EstradaAinda não há avaliações
- Introducción A La EmbriologíaDocumento36 páginasIntroducción A La EmbriologíaChristopher Barrionuevo GuevaraAinda não há avaliações
- Guía didáctica sobre función genética y herencia a partir de la estructura del ADNDocumento5 páginasGuía didáctica sobre función genética y herencia a partir de la estructura del ADNJose luis Lopez sanchezAinda não há avaliações
- Traduccion y Sintesis de ProteinasDocumento36 páginasTraduccion y Sintesis de ProteinasSara María Vargas CastilloAinda não há avaliações
- Transmisión de SimplesDocumento8 páginasTransmisión de SimplesDruman Vélez100% (1)
- Cerveza de Argentina - ¿Cómo Se Reproducen Las LevadurasDocumento2 páginasCerveza de Argentina - ¿Cómo Se Reproducen Las LevadurasJuan DornaAinda não há avaliações
- Homo Sapiens SapiensDocumento20 páginasHomo Sapiens SapiensElizaHGAinda não há avaliações
- Herencia MendelianaDocumento16 páginasHerencia MendelianaSaliza de Castro100% (1)
- Etapa 3 - Cuestionario Unidad 2 - Realizar Cuestionario PDFDocumento3 páginasEtapa 3 - Cuestionario Unidad 2 - Realizar Cuestionario PDFyuly barraganAinda não há avaliações